http://skepticwiki.org/index.php/Archaeopteryx#The_skull
"In modern birds, the bones of the pelvis and the lumbar, sacral, and a few caudal vertebrae are fused into a single, solid structure called the synsacrum. In Archaeopteryx, as in dinosaurs, the vertebrae are free and mobile."
Monday, May 31, 2010
* Cervical ribs
http://en.wikipedia.org/wiki/Cervical_vertebrae
"In many species, though not in mammals, the cervical vertebrae bear ribs. In many other groups, such as lizards and saurischian dinosaurs, the cervical ribs are large; in birds they are small and completely fused to the vertebrae. "
"In many species, though not in mammals, the cervical vertebrae bear ribs. In many other groups, such as lizards and saurischian dinosaurs, the cervical ribs are large; in birds they are small and completely fused to the vertebrae. "
Attaching the cervical vertebrae to the skull
http://skepticwiki.org/index.php/Archaeopteryx#The_skull
"Note how in Archaeopteryx the attachment of the cervical vertebrae to the skull is at the back of the head as in dinosaurs, not to the base of the skull as is found in modern birds."
Hypapophysis
http://scienceblogs.com/tetrapodzoology/2011/05/25/tiny-cretaceous-theropod/
Supplementary information
"Note how in Archaeopteryx the attachment of the cervical vertebrae to the skull is at the back of the head as in dinosaurs, not to the base of the skull as is found in modern birds."
Hypapophysis
http://scienceblogs.com/tetrapodzoology/2011/05/25/tiny-cretaceous-theropod/
Before discussing size further, let me note that the maniraptoran identification is pretty sound. On first seeing the vertebra, I was struck by the X-like shape of the neural arch (as seen in dorsal view), the large hypapophysis (a prominent keel located on the midline of the ventral surface) and the presence of fossae on the sides of the neural arch. It looked immediately like a maniraptoran cervical vertebra, especially that of an oviraptorosaur. This was confirmed by other details, like the shapes of the articular surfaces, the positions of the parapophyses, the shapes of the zygapophyses and so on. The presence of a rather large hypapophysis, combined with the position of the parapophyses (low down on the centrum), shows that the Ashdown maniraptoran vertebra is a posterior cervical: that is, a vertebra from near the base of the neck.http://rspb.royalsocietypublishing.org/content/early/2009/10/12/rspb.2009.1603
"In sharp contrast to the derived morphology of the skull and neck, the tail of Darwinopterus is identical to that of most basal clades, consisting of more than 20 caudals which, apart from the first three or four vertebrae, are long and rod-like and enclosed by a sheath of bony filaments composed of highly elongated ossified extensions of the zygapophyses and hypapophyses (figure 2e) (Wellnhofer 1975, 1978)."https://books.google.ca/books?id=YhAYWfPkJkQC&pg=PA68&lpg=PA68&dq=large+hypapophyses&source=bl&ots=V01o0idA18&sig=q5wnYf76h-IUDiP1sBpct8KGXOc&hl=en&sa=X&ei=Tf1gVYekFJWlyASk2IPQBg&ved=0CB8Q6AEwAA#v=onepage&q=large%20hypapophyses&f=false
They [hypapophyses] have not been reported in Archaeopteryx or any of the dinosaurs proposed as ancestors of the birds, but they appear in some fossils of modern birds.
We can only guess at the respiratory system of dinosaurs, and the appearance of well-developed thoracic hypapophyses would support the notion of something birdlike. None have been reported, however.
http://www.nature.com/nature/journal/v461/n7264/pdf/nature08322.pdf
Supplementary information
Unambiguous synapomorphies for major coelurosaurian clades:
Maniraptora: 46.1, 54.1, 57.1, 102.1, 105.1, 158.1, 161.0, 173.1, 175.1, and 263.1.
102. Anterior trunk vertebrae without prominent hypapophyses (0) or with large hypapophyses (1).
Sunday, May 30, 2010
Sternal Ribs
Another characteristic shared by pterosaurs and birds:
http://www.ploscollections.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0004497;jsessionid=A1CA60013B42AE8DCA4EDCACEF3A2BF7
The morphology of the trunk of pterosaurs differs from previous descriptions in several aspects that are crucial to lung ventilation and respiratory efficiency. Contrary to earlier reports [1], [7], [29], [30], pterosaur sternal ribs are not of uniform length and posterior elements commonly exhibit a two-fold or greater increase in length (Figs. 2, S1; Table S2). Consequently, and unlike recent reconstructions of pterosaurs which tend to show a horizontal or even posterodorsally sloping sternum, the posterior margin of the pterosaur sternum sloped posteroventrally, similar to birds[21]. (Leon P. A. M. Claessens1*, Patrick M. O'Connor2, David M. Unwin3)
http://www.ploscollections.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0004497;jsessionid=A1CA60013B42AE8DCA4EDCACEF3A2BF7
The morphology of the trunk of pterosaurs differs from previous descriptions in several aspects that are crucial to lung ventilation and respiratory efficiency. Contrary to earlier reports [1], [7], [29], [30], pterosaur sternal ribs are not of uniform length and posterior elements commonly exhibit a two-fold or greater increase in length (Figs. 2, S1; Table S2). Consequently, and unlike recent reconstructions of pterosaurs which tend to show a horizontal or even posterodorsally sloping sternum, the posterior margin of the pterosaur sternum sloped posteroventrally, similar to birds[21]. (Leon P. A. M. Claessens1*, Patrick M. O'Connor2, David M. Unwin3)
Aspiration Pump
Another characteristic shared by pterosaurs and birds:
http://www.ploscollections.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0004497;jsessionid=A1CA60013B42AE8DCA4EDCACEF3A2BF7
"The aspiration pump of pterosaurs maximised trunk expansion in the ventrocaudal region, while at the same time limiting the degrees of freedom of movement of the trunk in other directions. This provided greater control over the location, amount and timing of trunk expansion, thereby enabling precisely-timed localized generation of pressure gradients within the pulmonary system, a trait that is also present in living birds where it is of paramount importance for the generation of air flow patterns in the lungs [27], [36]." (Leon P. A. M. Claessens1*, Patrick M. O'Connor2, David M. Unwin3)
http://www.ploscollections.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0004497;jsessionid=A1CA60013B42AE8DCA4EDCACEF3A2BF7
"The aspiration pump of pterosaurs maximised trunk expansion in the ventrocaudal region, while at the same time limiting the degrees of freedom of movement of the trunk in other directions. This provided greater control over the location, amount and timing of trunk expansion, thereby enabling precisely-timed localized generation of pressure gradients within the pulmonary system, a trait that is also present in living birds where it is of paramount importance for the generation of air flow patterns in the lungs [27], [36]." (Leon P. A. M. Claessens1*, Patrick M. O'Connor2, David M. Unwin3)
Saturday, May 29, 2010
What is this?
Is this a pterosaur? Is this a modern bird ("neornithes")? Is this a maniraptor dinosaur?

How about this one:

And this one?

And this one?

Which picture does not go with the others?

How about this one:

And this one?

And this one?

Which picture does not go with the others?
Dino to bird series based on "hypothetical animal"
After all is said and done and all the charts and relationships are analyzed here is what the dino to bird theory comes down to:
http://en.wikipedia.org/wiki/Eumaniraptora
"The name Paraves was coined by Paul Sereno in 1997.[1] The clade was defined by Sereno in 1998 as a branch-based clade containing all Maniraptora closer to Neornithes (which includes all the birds living in the world today) than to Oviraptor.[2]
The ancestral paravian is a hypothetical animal; the first common ancestor of birds, dromaeosaurids, and troodontids which was not also ancestral to oviraptorosaurs. Little can be said with certainty about this animal. The work of Turner et al. (2007) suggested that the ancestral paravian could not glide or fly, and that it was most likely small (around 65 centimeters long and 600–700 grams in mass).[3] But the work of Xu et al. (2003), (2005) and Hu et al (2009) provide examples of basal and early paravians with four wings, including members of the Avialae (Pedopenna), Dromaeosauridae (Microraptor), and Troodontidae (Anchiornis).[4][5][6]"
http://en.wikipedia.org/wiki/Eumaniraptora
"The name Paraves was coined by Paul Sereno in 1997.[1] The clade was defined by Sereno in 1998 as a branch-based clade containing all Maniraptora closer to Neornithes (which includes all the birds living in the world today) than to Oviraptor.[2]
The ancestral paravian is a hypothetical animal; the first common ancestor of birds, dromaeosaurids, and troodontids which was not also ancestral to oviraptorosaurs. Little can be said with certainty about this animal. The work of Turner et al. (2007) suggested that the ancestral paravian could not glide or fly, and that it was most likely small (around 65 centimeters long and 600–700 grams in mass).[3] But the work of Xu et al. (2003), (2005) and Hu et al (2009) provide examples of basal and early paravians with four wings, including members of the Avialae (Pedopenna), Dromaeosauridae (Microraptor), and Troodontidae (Anchiornis).[4][5][6]"
Friday, May 28, 2010
Another thing to watch for
Let's take a look at another form of misleading dino to bird argument. Evidence which can be explained in a very simple alternate way is instead touted to support a dino to bird theory.
As an example, here is an article on the subject of dinosaur "air sacs" and the idea that dinosaurs had a breathing system like modern birds:
http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0003303
http://en.wikipedia.org/wiki/Aerosteon
First the researchers simply found "pneumatization of skeletal bone, including pneumatic hollowing of the furcula and ilium."
Everything else - about air sacs and breathing systems - is simply speculation.
But this speculation is then simply passed on as given, as we see in the wikipedia reference.
And in fact the simpler explanations (that are mentioned at the end) - for example "thermal regulation" - are ignored.
This is something we must keep alert to. Evidence which can be explained in a very simple alternate way is instead touted to support a dino to bird theory.
When the dino to bird theorist states that there is a huge amount of supporting evidence, it is often just this kind of speculation that is actually at the bottom of it.
Additional irony:
Even Sereno is "especially intrigued" by the simpler, reasonable explanation:
http://newswise.com/articles/view/544722/
Evidence for Avian Intrathoracic Air Sacs in a New Predatory Dinosaur from Argentina
http://rspb.royalsocietypublishing.org/content/275/1631/157.full.pdf
As an example, here is an article on the subject of dinosaur "air sacs" and the idea that dinosaurs had a breathing system like modern birds:
http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0003303
Evidence for Avian Intrathoracic Air Sacs in a New Predatory Dinosaur from Argentina
"Living birds possess a unique heterogeneous pulmonary system composed of a rigid, dorsally-anchored lung and several compliant air sacs that operate as bellows, driving inspired air through the lung [during exhalation]. Evidence from the fossil record for the origin and evolution of this system is extremely limited, because lungs do not fossilize and because the bellow-like air sacs in living birds only rarely penetrate (pneumatize) skeletal bone and thus leave a record of their presence.
Methodology/Principal Findings
We describe a new predatory dinosaur from Upper Cretaceous rocks in Argentina, Aerosteon riocoloradensis gen. et sp. nov., that exhibits extreme pneumatization of skeletal bone, including pneumatic hollowing of the furcula and ilium. In living birds, these two bones are pneumatized by diverticulae of air sacs (clavicular, abdominal) that are involved in pulmonary ventilation. We also describe several pneumatized gastralia (“stomach ribs”), which suggest that diverticulae of the air sac system were present in surface tissues of the thorax.
Conclusions/SignificanceWe present a four-phase model for the evolution of avian air sacs and costosternal-driven lung ventilation based on the known fossil record of theropod dinosaurs and osteological correlates in extant birds:(1) Phase I—Elaboration of paraxial cervical air sacs in basal theropods no later than the earliest Late Triassic.(2) Phase II—Differentiation of avian ventilatory air sacs, including both cranial (clavicular air sac) and caudal (abdominal air sac) divisions, in basal tetanurans during the Jurassic. A heterogeneous respiratory tract with compliant air sacs, in turn, suggests the presence of rigid, dorsally attached lungs with flow-through ventilation.(3) Phase III—Evolution of a primitive costosternal pump in maniraptoriform theropods before the close of the Jurassic.(4) Phase IV—Evolution of an advanced costosternal pump in maniraptoran theropods before the close of the Jurassic.In addition, we conclude:(5) The advent of avian unidirectional lung ventilation is not possible to pinpoint, as osteological correlates have yet to be identified for uni- or bidirectional lung ventilation.(6) The origin and evolution of avian air sacs may have been driven by one or more of the following three factors: flow-through lung ventilation, locomotory balance, and/or thermal regulation."Paul C. Sereno1*, Ricardo N. Martinez2, Jeffrey A. Wilson3, David J. Varricchio4, Oscar A. Alcober2, Hans C. E. Larsson5
http://en.wikipedia.org/wiki/Aerosteon
Aerosteon is a genus of allosauroid theropod dinosaur from the Late Cretaceous period of Argentina. Its remains were discovered in 1996 in the province of Mendoza. They show evidence of a bird-like respiratory system.[1]
Sereno theorises that this respiratory system may have developed to assist with regulating body temperature and was later co-opted for breathing.[1]Note a few things here.
First the researchers simply found "pneumatization of skeletal bone, including pneumatic hollowing of the furcula and ilium."
Everything else - about air sacs and breathing systems - is simply speculation.
But this speculation is then simply passed on as given, as we see in the wikipedia reference.
And in fact the simpler explanations (that are mentioned at the end) - for example "thermal regulation" - are ignored.
This is something we must keep alert to. Evidence which can be explained in a very simple alternate way is instead touted to support a dino to bird theory.
When the dino to bird theorist states that there is a huge amount of supporting evidence, it is often just this kind of speculation that is actually at the bottom of it.
Additional irony:
Even Sereno is "especially intrigued" by the simpler, reasonable explanation:
http://newswise.com/articles/view/544722/
Sereno, a National Geographic Explorer-in-Residence, said he is especially intrigued by heat loss, given that Aerosteon was likely a high-energy predator with feathers but without the sweat glands that birds possess. At approximately 30 feet in length and weighing as much as an elephant, Aerosteon might well have used an air system under the skin to rid itself of unwanted heat.http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0003303
Evidence for Avian Intrathoracic Air Sacs in a New Predatory Dinosaur from Argentina
Paul C. Sereno et al
Research on the gastral cuirass in archosaurs led to the suggestion that it may have functioned as an accessory aspiration pump in nonavian dinosaurs [81]–[83]. Although Claessens drew attention to the relationship between the gastral cuirass and abdominal air sacs, he concluded that “it appears impossible to ascertain exactly when lung diverticula stretching throughout the whole body cavity or unidirectional airflow originated” [83:102]. Later a “caudal origin model” for air sacs and flow-through lung ventilation (either uni- or bidirectional) was proposed [33: 255] based on (1) the presence of abdominal air sacs (inferred from posterior dorsal and sacral pneumaticity), (2) a dynamic gastral cuirass, and (3) vertebrocostal articulations in the posterior ribcage that allow greater excursion during aspiration (inferred from the more horizontal arrangement of posterior rib articulations). An independent study of rib morphology, in contrast, concluded that nonavian dinosaurs were characterized by an “anteriorly ventilated bellows lung”[84: 47].
In 1868 Thomas Huxley first proposed that dinosaurs were the direct ancestors of birds and subsequent analyses have identified a suite of ‘avian’ characteristics in theropod dinosaurs. Ossified uncinate processes are found in most species of extant birds and also occur in extinct non-avian maniraptoran dinosaurs. Their presence in these dinosaurs represents another morphological character linking them to Aves, and further
supports the presence of an avian-like air-sac respiratory system in theropod dinosaurs, prior to the evolution of flight. Here we report a phylogenetic analysis of the presence of uncinate processes in Aves and non-avian maniraptoran dinosaurs indicating that these were homologous structures. Furthermore, recent work on Canada geese has demonstrated that uncinate processes are integral to the mechanics of avian ventilation,
facilitating both inspiration and expiration. In extant birds, uncinate processes function to increase the mechanical advantage for movements of the ribs and sternum during respiration. Our study presents a mechanism whereby uncinate processes, in conjunction with lateral and ventral movements of the sternum and gastral basket, affected avian-like breathing mechanics in extinct non-avian maniraptoran dinosaurs.
Pneumatic (air-filled) postcranial bones are unique to birds among extant tetrapods. Unambiguous skeletal correlates of postcranial pneumaticity first appeared in the Late Triassic (approximately 210 million years ago), when they evolved independently in several groups of bird-line archosaurs (ornithodirans). These include the theropod dinosaurs (of which birds are extant representatives), the pterosaurs, and sauropodomorph dinosaurs. Postulated functions of skeletal pneumatisation include weight reduction in large-bodied or flying taxa, and density reduction resulting in
energetic savings during foraging and locomotion. However, the influence of these hypotheses on the early evolution of pneumaticity has not been studied in detail previously. We review recent work on the significance of pneumaticity for understanding the biology of extinct ornithodirans, and present detailed new data on the proportion of the skeleton that was pneumatised in 131 non-avian theropods and Archaeopteryx. This includes all taxa known from significant postcranial remains. Pneumaticity of the cervical and anterior dorsal vertebrae occurred early in theropod evolution.
This ‘common pattern’ was conserved on the line leading to birds, and is likely present in Archaeopteryx. Increases in skeletal pneumaticity occurred independently in as many as 12 lineages, highlighting a remarkably high number of parallel acquisitions of a bird-like feature among non-avian theropods. Using a quantitative comparative framework, we show that evolutionary increases in skeletal pneumaticity are significantly concentrated in lineages with large body size, suggesting that mass reduction in response to gravitational constraints at large body sizes influenced the early evolution of pneumaticity. However, the body size threshold for extensive pneumatisation is lower in theropod lineages more closely related to birds (maniraptorans). Thus, relaxation of the relationship between body size and pneumatisation preceded the origin of birds and cannot be explained as an adaptation for flight. We hypothesise that skeletal density modulation in small, non-volant, maniraptorans resulted in energetic savings as part of a multi-system response to increased metabolic demands. Acquisition of extensive postcranial pneumaticity in small-bodied maniraptorans may indicate avian-like high-performance endothermy.
http://college.holycross.edu/faculty/lclaesse/Claessens_etal_2009_PLoS.pdf
https://en.wikipedia.org/wiki/Air_sacs
Pterosaurs, enigmatic extinct Mesozoic reptiles, were the first vertebrates to achieve true flapping flight. Various lines of evidence provide strong support for highly efficient wing design, control, and flight capabilities. However, little is known of the pulmonary system that powered flight in pterosaurs. We investigated the structure and function of the pterosaurian breathing apparatus through a broad scale comparative study of respiratory structure and function in living and extinct archosaurs, using computer-assisted tomographic (CT) scanning of pterosaur and bird skeletal remains, cineradiographic (Xray film) studies of the skeletal breathing pump in extant birds and alligators, and study of skeletal structure in historic fossil specimens. In this report we present various lines of skeletal evidence that indicate that pterosaurs had a highly effective flow-through respiratory system, capable of sustaining powered flight, predating the appearance of an analogous breathing system in birds by approximately seventy million years. Convergent evolution of gigantism in several Cretaceous pterosaur lineages was made possible through body density reduction by expansion of the pulmonary air sac system throughout the trunk and the distal limb girdle skeleton, highlighting the importance of respiratory adaptations in pterosaur evolution, and the dramatic effect of the release of physical constraints on morphological diversification and evolutionary radiation.http://en.wikipedia.org/wiki/Mirischia
The specimen is also unusual in that it preserves some soft tissue remains: apart from the intestine, what the describers interpreted to have been an air sac was preserved between its pubic and ischial bones in the form of a vacuity. Previous workers had suggested that non-avian theropods might — like birds — possess post-cranial air sacs, and Mirischia seems to confirm that. Another notable trait is the exceptional thinness of the bone wall of all skeletal elements.[2]
https://en.wikipedia.org/wiki/Air_sacs
John Ruben et al. (1997, 1999, 2003, 2004) disputed this and suggested that dinosaurs had a "tidal" respiratory system (in and out) powered by a crocodile-like hepatic piston mechanism – muscles attached mainly to the pubis pull the liver backwards, which makes the lungs expand to inhale; when these muscles relax, the lungs return to their previous size and shape, and the animal exhales. They also presented this as a reason for doubting that birds descended from dinosaurs.[5][6][7][8][9]Critics have claimed that, without avian air sacs, modest improvements in a few aspects of a modern reptile's circulatory and respiratory systems would enable the reptile to achieve 50% to 70% of the oxygen flow of a mammal of similar size,[10] and that lack of avian air sacs would not prevent the development of endothermy.[11] Very few formal rebuttals have been published in scientific journals of Ruben et al.’s claim that dinosaurs could not have had avian-style air sacs; but one points out that the Sinosauropteryx fossil on which they based much of their argument was severely flattened and therefore it was impossible to tell whether the liver was the right shape to act as part of a hepatic piston mechanism.[12] Some recent papers simply note without further comment that Ruben et al. argued against the presence of air sacs in dinosaurs.[13]
Thursday, May 27, 2010
More on the brain
Pterosaurs are like modern birds right down to the details:
http://digimorph.org/specimens/Anhanguera_santanae/Witmer_02048.pdf
"We use the avian equation to estimate the pterosaur EQs because pterosaur brains filled the endocranial cavity (as in birds, but unlike in reptiles)."
(Lawrence M. Witmer1, Sankar Chatterjee2, Jonathan Franzosa3 & Timothy Rowe3)
http://digimorph.org/specimens/Anhanguera_santanae/Witmer_02048.pdf
"We use the avian equation to estimate the pterosaur EQs because pterosaur brains filled the endocranial cavity (as in birds, but unlike in reptiles)."
(Lawrence M. Witmer1, Sankar Chatterjee2, Jonathan Franzosa3 & Timothy Rowe3)
Neuroanatomy (3)
This is an excellent source of info about pterosaur and modern bird similarities:
http://digimorph.org/specimens/Anhanguera_santanae/Witmer_02048.pdf
"Virtual endocasts derived from CT scans of nearly complete skulls of two pterosaurs (Fig. 1)—the more basal Rhamphorhynchus and the pterodactyloid Anhanguera—are the most complete to date.
They confirm some previous findings of birdlike attributes1–8: expansion of the cerebrum and cerebellum, displacing the enlarged optic tecta (lobes) ventrolaterally; small olfactory areas; and enlarged flocculi (cerebellar auricles) (Fig. 2). Despite these structural similarities, the brains of Rhamphorhynchus and Anhanguera, relative to body mass, do not fall [quite] within the range of extant birds, although they were enlarged relative to extant nonavian reptiles4,5,11 (Fig. 3; see Methods). Moreover, comparisons of total brain mass do not reveal differences in relative size of brain components (and hence underlying neural organization). For example, the enormous flocculi of pterosaurs probably outweighed the optic tecta, whereas the reverse is certainly true in birds.
Nevertheless, pterosaurs do possess a number of avian neuroanatomical traits that may well be associated with the sensory and coordination functions necessary for flight. Jerison4 suggested that avian brains were relatively larger than those of pterosaurs because birds evolved in the environmentally complex and neurologically challenging arboreal habitat that required greater neural processing and hence greater mass. That may be true, but another factor is that birds and pterosaurs had different phylogenetic starting points: pterosaurs evolved from relatively very smallbrained basal archosaurs, whereas birds evolved from theropod dinosaurs that had already initiated a substantial trend of brain expansion5,9. The virtual endocasts include the semicircular canals (Fig. 2), which had previously been only partially known for one pterosaur, Parapsicephalus1. The entire osseous labyrinth is preserved bilaterally in Anhanguera and the large majority of it is preserved in Rhamphorhynchus. The semicircular canal system is greatly expanded, with the long canals encircling the flocculus. Its general arrangement closely resembles that of birds and some other dinosaurs5, but, whereas it is relatively modest in these groups, the vestibular apparatus is relatively much larger in the pterosaurs." (Lawrence M. Witmer1, Sankar Chatterjee2, Jonathan Franzosa3 & Timothy Rowe3)
Note:
The article says:"the brains of Rhamphorhynchus and Anhanguera, relative to body mass, do not fall [quite] within the range of extant birds."
I have inserted the word "quite" because you must look at Figure 3 of the report to appreciate how very close the pterosaur is to being within the range of extant birds. And also review the Methods section to see the roughness of the calculation.
Also keep in mind that I am not claiming that pterosaurs were identical to modern birds. There was obviously a development of the brain in the transition from pterosaur to modern bird. But not much change was required (if any) in this respect.
http://digimorph.org/specimens/Anhanguera_santanae/Witmer_02048.pdf
"Virtual endocasts derived from CT scans of nearly complete skulls of two pterosaurs (Fig. 1)—the more basal Rhamphorhynchus and the pterodactyloid Anhanguera—are the most complete to date.
They confirm some previous findings of birdlike attributes1–8: expansion of the cerebrum and cerebellum, displacing the enlarged optic tecta (lobes) ventrolaterally; small olfactory areas; and enlarged flocculi (cerebellar auricles) (Fig. 2). Despite these structural similarities, the brains of Rhamphorhynchus and Anhanguera, relative to body mass, do not fall [quite] within the range of extant birds, although they were enlarged relative to extant nonavian reptiles4,5,11 (Fig. 3; see Methods). Moreover, comparisons of total brain mass do not reveal differences in relative size of brain components (and hence underlying neural organization). For example, the enormous flocculi of pterosaurs probably outweighed the optic tecta, whereas the reverse is certainly true in birds.
Nevertheless, pterosaurs do possess a number of avian neuroanatomical traits that may well be associated with the sensory and coordination functions necessary for flight. Jerison4 suggested that avian brains were relatively larger than those of pterosaurs because birds evolved in the environmentally complex and neurologically challenging arboreal habitat that required greater neural processing and hence greater mass. That may be true, but another factor is that birds and pterosaurs had different phylogenetic starting points: pterosaurs evolved from relatively very smallbrained basal archosaurs, whereas birds evolved from theropod dinosaurs that had already initiated a substantial trend of brain expansion5,9. The virtual endocasts include the semicircular canals (Fig. 2), which had previously been only partially known for one pterosaur, Parapsicephalus1. The entire osseous labyrinth is preserved bilaterally in Anhanguera and the large majority of it is preserved in Rhamphorhynchus. The semicircular canal system is greatly expanded, with the long canals encircling the flocculus. Its general arrangement closely resembles that of birds and some other dinosaurs5, but, whereas it is relatively modest in these groups, the vestibular apparatus is relatively much larger in the pterosaurs." (Lawrence M. Witmer1, Sankar Chatterjee2, Jonathan Franzosa3 & Timothy Rowe3)
Note:
The article says:"the brains of Rhamphorhynchus and Anhanguera, relative to body mass, do not fall [quite] within the range of extant birds."
I have inserted the word "quite" because you must look at Figure 3 of the report to appreciate how very close the pterosaur is to being within the range of extant birds. And also review the Methods section to see the roughness of the calculation.
Also keep in mind that I am not claiming that pterosaurs were identical to modern birds. There was obviously a development of the brain in the transition from pterosaur to modern bird. But not much change was required (if any) in this respect.
Neuroanatomy (2)
Dinosaur:
Incisivosaurus gauthieri, Oviraptorid
http://digimorph.org/specimens/Incisivosaurus_gauthieri/
Dr. Xing Xu, Dr. Yoshimura Kobayashi, Dr. Yusuke Matsufune, and Dr. Mark A. Norell

"The imagery on this page is the basis for a paper entitled Cranial osteology of the theropod dinosaur Incisivosaurus gauthieri (Theropoda: Oviraptorosauria), by A.M. Balanoff, X. Xu, Y. Kobayashi, Y. Matsufune, and M.A. Norell, 2009 (American Museum Novitates 3651:1-35)."
Incisivosaurus gauthieri, Oviraptorid
http://digimorph.org/specimens/Incisivosaurus_gauthieri/
Dr. Xing Xu, Dr. Yoshimura Kobayashi, Dr. Yusuke Matsufune, and Dr. Mark A. Norell
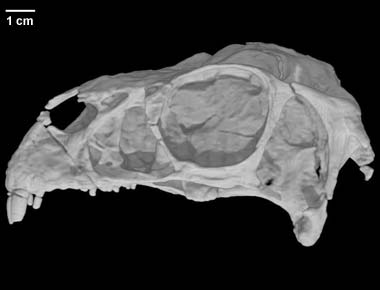
"The imagery on this page is the basis for a paper entitled Cranial osteology of the theropod dinosaur Incisivosaurus gauthieri (Theropoda: Oviraptorosauria), by A.M. Balanoff, X. Xu, Y. Kobayashi, Y. Matsufune, and M.A. Norell, 2009 (American Museum Novitates 3651:1-35)."
We provide a description of the holotype skull of the unusual oviraptorosaur Incisivosaurus gauthieri. Previous phylogenetic analyses have placed this taxon firmly within Oviraptorosauria near the base of the clade; however, until now only a cursory description of this important specimen was available. The presence of many primitive characteristics (e.g., maxillary and dentary teeth as well as an extended palate and rostrum) indicates that the observed similarities between avians and derived oviraptorids are convergences rather than shared derived characters. In addition, we clarify previous descriptions of several ambiguous anatomical features, most notably of the palate. We also employ a computed tomographic (CT) analysis, which allows for a more complete description of the braincase and the reconstruction of an endocranial endocast. CT imagery reveals features that were before unobtainable, such as the presence of a replacement tooth behind the large rodentiform incisor in the premaxilla. This arrangement indicates that although the incisiform teeth of I. gauthieri are morphologically distinct they are replaced in typical archosaurian fashion.The simpler explanation is that oviraptors are secondarily flightless members of paraves.
Neuroanatomy (1)
Notice the neuroanatomical differences between the primitive pterosaur (Rhamphorhynchus muensteri) and the advanced pterosaur (Anhanguera santanae).
Anhanguera santanae , Pterosaur
, Pterosaur
http://digimorph.org/specimens/Anhanguera_santanae/
Ohio University College of Osteopathic Medicine

Rhamphorhynchus muensteri, Pterosaur
http://digimorph.org/specimens/Rhamphorhynchus_muensteri/
Ohio University College of Osteopathic Medicine

Anhanguera santanae
, Pterosaurhttp://digimorph.org/specimens/Anhanguera_santanae/
Ohio University College of Osteopathic Medicine
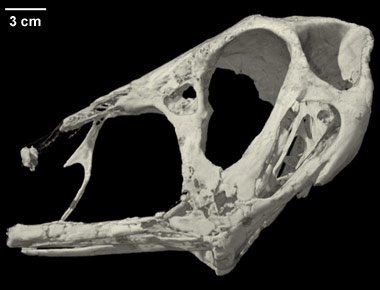
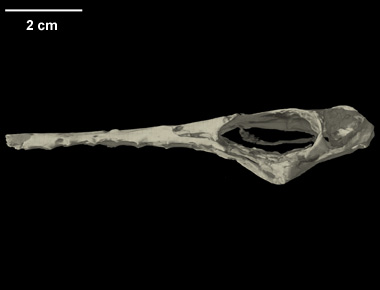
Neuroanatomy of flying reptiles and implications for flight, posture and behaviour
Lawrence M. Witmer, Sankar Chatterjee, Jonathan Franzosa & Timothy Rowe
| ABSTRACT: "Comparison of birds and pterosaurs, the two archosaurian flyers, sheds light on adaptation to an aerial lifestyle. The neurological basis of control holds particular interest in that flight demands on sensory integration, equilibrium, and muscular coordination are acute. Here we compare the brain and vestibular apparatus in two pterosaurs [Rhamphorhynchus muensteri and Anhanguera santanae] based on high-resolution computed tomographic (CT) scans from which we constructed digital endocasts. Although general neural organization resembles birds, pterosaurs had smaller brains relative to body mass than do birds. This difference probably has more to do with phylogeny than flight, in that birds evolved from nonavian theropods that had already established trends for greater encephalization. Orientation of the osseous labyrinth relative to the long axis of the skull was also different in these two species, suggesting very different head postures and reflecting differing behaviours. Their enlarged semicircular canals reflect a highly refined organ of equilibrium, which is concordant with pterosaurs being visually based, aerial predators. Their enormous cerebellar floccular lobes may suggest neural integration of extensive sensory information from the wing, further enhancing eye- and neck-based reflex mechanisms for stabilizing gaze." |

 , Pterosaur
, Pterosaurhttp://digimorph.org/specimens/Rhamphorhynchus_muensteri/
Ohio University College of Osteopathic Medicine
|

Wednesday, May 26, 2010
Starting to match them up
Now that we have seen the 4 major categories of modern birds (Neoaves, Anseriformes, Galliformes, and Palaeognathae) - let's look at the categories of advanced pterosaurs:
ADVANCED PTEROSAURS (Pterodactyloidea)
Superfamily Ornithocheiroidea
VULTURE
-------------------------------------------------
ALBATROSS
-----------------------------------------------
PELICAN
----------------------------------------------
FLAMINGO
-------------------------------------------
STORK
ADVANCED PTEROSAURS (Pterodactyloidea)
Superfamily Ornithocheiroidea
- Family Istiodactylidae
- Family Boreopteridae[2]
- Family Ornithocheiridae
- Family Pteranodontidae
- Superfamily Dsungaripteroidea
- Superfamily Ctenochasmatoidea
- Cycnorhamphus
- Feilongus
- Pterodactylus (PELICAN?/Neoaves)
- Family Ctenochasmatidae
- Elanodactylus[1]
- Gegepterus[2]
- Subfamily Ctenochasmatinae
- Subfamily Gnathosaurinae
- Cearadactylus
- Gnathosaurus
- Huanhepterus
- Plataleorhynchus
- "Pterodactylus" longicollum
- Superfamily Azhdarchoidea
- Family Chaoyangopteridae[1]
- Family Lonchodectidae
- Family Tapejaridae
- Subfamily Tapejarinae
- Subfamily Thalassodrominae [3]
- Family Azhdarchidae (STORK?/Neoaves)
- Possible valid azhdarchid genera
- Bennettazhia
- Bogolubovia
- Doratorhynchus
- "Ornithocheirus" bunzeli
VULTURE
| Class: | Aves |
| Families | |
|---|---|
-------------------------------------------------
ALBATROSS
| Neoaves | |
| Order: | Procellariiformes |
| Family: | Diomedeidae G.R. Gray 1840[1] |
PELICAN
| Aves Infraclass: Neognathae Neoaves | |
| Order: | Pelecaniformes |
| Family: | Pelecanidae Rafinesque, 1815 |
| Genus: | Pelecanus |
FLAMINGO
| Aves | |
| Infraclass: | Neognathae Neoaves |
| Order: | Phoenicopteriformes Fürbringer, 1888 |
| Family: | Phoenicopteridae Bonaparte, 1831 |
| Genus: | Phoenicopterus Phoenicoparrus |
STORK
| Neornithes | |
| Infraclass: | Neognathae |
| Superorder: | Neoaves |
| Order: | Ciconiiformes |
| Family: | Ciconiidae Gray, 1840 |
The transition to modern birds
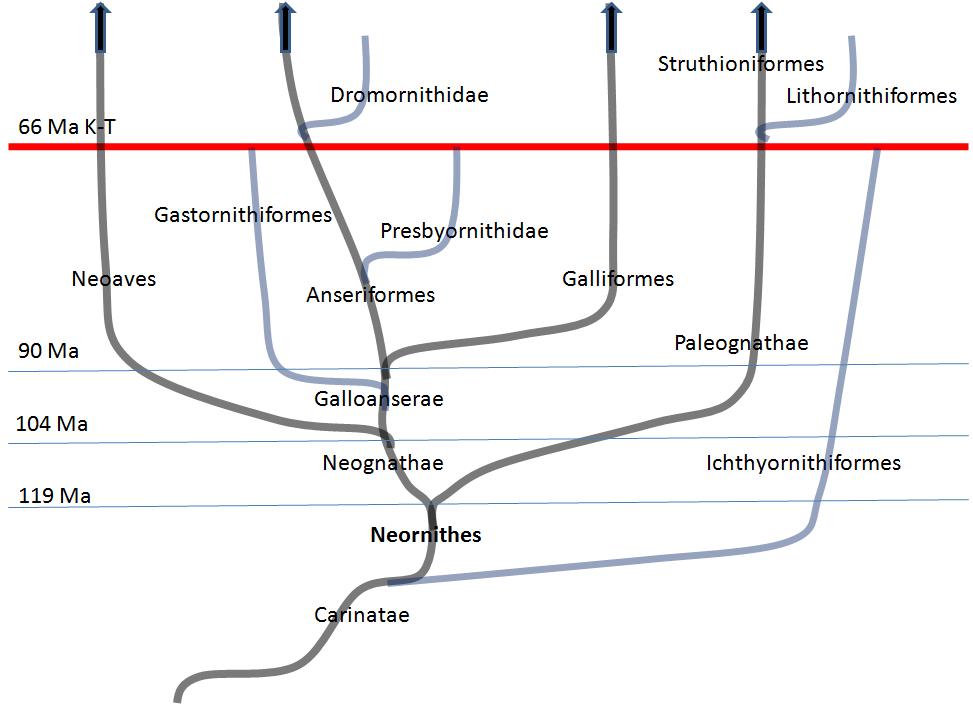
Here is a chart outlining the current thinking about the most recent evolution phase of modern birds.

Consider the idea that pterosaur types developed into their corresponding modern bird types (within Neoaves, Anseriformes, and Galliformes).
Palaeognathae requires more research.
http://en.wikipedia.org/wiki/Neornithes
"Modern birds (subclass Neornithes) are the most recent common ancestor of all living birds (class Aves) and all its descendants.
Consider the idea that pterosaur types developed into their corresponding modern bird types (within Neoaves, Anseriformes, and Galliformes).
Palaeognathae requires more research.
http://en.wikipedia.org/wiki/Neornithes
"Modern birds (subclass Neornithes) are the most recent common ancestor of all living birds (class Aves) and all its descendants.
Modern birds are characterised by feathers, a beak with no teeth (ancient birds had teeth), the laying of hard-shelled eggs, a high metabolic rate, a four-chambered heart, and a lightweight but strong skeleton. All birds have forelimbs modified as wings and most can fly, with some exceptions including ratites, penguins, and a number of diverse endemic island species. Birds also have unique digestive and respiratory systems that are highly adapted for flight. Some birds, especially corvids and parrots, are among the most intelligent animal species; a number of bird species have been observed manufacturing and using tools, and many social species exhibit cultural transmission of knowledge across generations."
Subclass Neornithes
- Palaeognathae:
- Struthioniformes, Ostrich, emus, kiwis, and allies
- Tinamiformes, tinamous
- Neognathae:
- Galloanserae:
- Anseriformes, waterfowl
- Galliformes, fowl
- Neoaves:
- Gaviiformes, loons
- Podicipediformes, grebes
- Procellariiformes, albatrosses, petrels, and allies
- Sphenisciformes, penguins
- Pelecaniformes, pelicans and allies
- Ciconiiformes, storks and allies
- Phoenicopteriformes, flamingos
- Falconiformes, falcons, eagles, hawks and allies
- Gruiformes, cranes and allies
- Charadriiformes, gulls, button-quail, plovers and allies
- Pteroclidiformes, sandgrouse
- Columbiformes, doves and pigeons
- Psittaciformes, parrots and allies
- Cuculiformes, cuckoos, turacos, hoatzin
- Strigiformes, owls
- Caprimulgiformes, nightjars and allies
- Apodiformes, swifts and hummingbirds
- Coraciiformes, kingfishers
- Piciformes, woodpeckers and allies
- Trogoniformes, trogons
- Coliiformes, mousebirds
- Passeriformes, passerines
- Galloanserae:
Tuesday, May 25, 2010
Skulls
http://en.wikipedia.org/wiki/Bird
"The skull bones [of modern birds] are fused and do not show cranial sutures.[39]"
(Frank (1995). Ornithology. )
http://en.wikipedia.org/wiki/Diapsid
"Diapsids ("two arches") are a group of reptiles that developed two holes (temporal fenestra) in each side of their skulls, about 300 million years ago during the late Carboniferous period. Living diapsids are extremely diverse, and include all crocodiles, lizards, snakes, and tuatara. Under modern classification systems, even birds are considered diapsids, since they evolved from diapsid ancestors and are nested within the diapsid clade. While some diapsids have lost either one hole (lizards), or both holes (snakes), or even have a heavily restructured skull (modern birds), they are still classified as diapsids based on their ancestry."
http://www.elp.manchester.ac.uk/pub_projects/2003/MNZO0MLK/lecture12.htm
"Note that the [modern] birds are clearly derived archosaurs, and in a phylogenetic classification should be included, but with their expanded brains and slight jaws (beaks), the diapsid nature of their skull has been lost"
http://archosaurmusings.wordpress.com/2008/06/15/pterosaur-origins-where-did-they-come-from/
"Thanks to the two posterior holes in the skull (the upper and lower temporal fenestrae for those in the know) we know pterosaurs are diapsid reptiles, but after this it gets really tricky, and the diapsids are a big group (it includes snakes, crocodile, dinosaurs, lizards, various marine reptiles, and all kinds of odds and sods like rhynchosaurs). Pterosaurs, rather obviously, are highly adapted for their flying lifestyle and as a result their anatomy has changed a huge amount. Not just the obviously elongate fourth finger, but the skull bones have kind of merged," (David Hone)
http://whenpigsfly-returns.blogspot.com/2007/08/pterosaurs-what-hell-are-they.html
"Also of note is that pterosaurs lack a mandibular fenestrae, a hole in the jaw that is also diagnostic of the Archosauriformes. If pterosaurs are archosauriforms, then that means that they, for whatever reason, secondarily lost the mandibular fenestrae. This makes no sense at all, given that pterosaurs excavated their skeletons like crazy in an effort to become more lightweight."
http://www.sciencemag.org/cgi/content/full/276/5318/1543
"This condition is highly variable among modern birds, which may show one, two, or no fenestrae (22). But modern birds differ from nonavian theropods in that the dentary lining presents most of the ventral margin of the rostral mandibular fenestra both laterally and medially"
"The bird also has a postorbital bone (Fig. 3), which articulates rostrodorsally to the frontal and caudally to the squamosal. This bone bears a ventral, splint-like jugal process that does not contact the jugal bar and only partially separates the orbit from the infratemporal fenestra. The postorbital bone is absent in modern birds but is present in nonavian theropods (19). Workers have argued about this bone's presence in Archaeopteryx (7-9, 15, 16), but the presence of a postorbital in the new fossil casts doubt on interpretations that it is absent in Archaeopteryx."
"Equally interesting is the presence of a tetraradiated squamosal that, in contrast to modern birds, is not incorporated into the braincase (Fig. 3). Its rostrolateral process terminates in a bifurcated facet for articulation with the postorbital."
http://icb.oxfordjournals.org/cgi/content/full/40/4/687
"The most important bird from the Liaoning deposits in China is Confuciusornis the only bird besides Archaeopteryx with enlarged claws for climbing. It also has a full set of gastralia and a postorbital bone that meets the jugal." (Larry D. Martin2,1 and Stephan A. Czerkas2 )
http://www.freethought-forum.com/forum/showthread.php?t=11592&garpg=3

"In the drawing, the “orbit” is the eye socket, “Sq” indicates the squamosal bone, one of the bones that makes up the skull, and “Po” indicates the postorbital bone."
"The skull bones [of modern birds] are fused and do not show cranial sutures.[39]"
(Frank (1995). Ornithology. )
http://en.wikipedia.org/wiki/Diapsid
"Diapsids ("two arches") are a group of reptiles that developed two holes (temporal fenestra) in each side of their skulls, about 300 million years ago during the late Carboniferous period. Living diapsids are extremely diverse, and include all crocodiles, lizards, snakes, and tuatara. Under modern classification systems, even birds are considered diapsids, since they evolved from diapsid ancestors and are nested within the diapsid clade. While some diapsids have lost either one hole (lizards), or both holes (snakes), or even have a heavily restructured skull (modern birds), they are still classified as diapsids based on their ancestry."
http://www.elp.manchester.ac.uk/pub_projects/2003/MNZO0MLK/lecture12.htm
"Note that the [modern] birds are clearly derived archosaurs, and in a phylogenetic classification should be included, but with their expanded brains and slight jaws (beaks), the diapsid nature of their skull has been lost"
http://archosaurmusings.wordpress.com/2008/06/15/pterosaur-origins-where-did-they-come-from/
"Thanks to the two posterior holes in the skull (the upper and lower temporal fenestrae for those in the know) we know pterosaurs are diapsid reptiles, but after this it gets really tricky, and the diapsids are a big group (it includes snakes, crocodile, dinosaurs, lizards, various marine reptiles, and all kinds of odds and sods like rhynchosaurs). Pterosaurs, rather obviously, are highly adapted for their flying lifestyle and as a result their anatomy has changed a huge amount. Not just the obviously elongate fourth finger, but the skull bones have kind of merged," (David Hone)
http://whenpigsfly-returns.blogspot.com/2007/08/pterosaurs-what-hell-are-they.html
"Also of note is that pterosaurs lack a mandibular fenestrae, a hole in the jaw that is also diagnostic of the Archosauriformes. If pterosaurs are archosauriforms, then that means that they, for whatever reason, secondarily lost the mandibular fenestrae. This makes no sense at all, given that pterosaurs excavated their skeletons like crazy in an effort to become more lightweight."
http://www.sciencemag.org/cgi/content/full/276/5318/1543
"This condition is highly variable among modern birds, which may show one, two, or no fenestrae (22). But modern birds differ from nonavian theropods in that the dentary lining presents most of the ventral margin of the rostral mandibular fenestra both laterally and medially"
"The bird also has a postorbital bone (Fig. 3), which articulates rostrodorsally to the frontal and caudally to the squamosal. This bone bears a ventral, splint-like jugal process that does not contact the jugal bar and only partially separates the orbit from the infratemporal fenestra. The postorbital bone is absent in modern birds but is present in nonavian theropods (19). Workers have argued about this bone's presence in Archaeopteryx (7-9, 15, 16), but the presence of a postorbital in the new fossil casts doubt on interpretations that it is absent in Archaeopteryx."
"Equally interesting is the presence of a tetraradiated squamosal that, in contrast to modern birds, is not incorporated into the braincase (Fig. 3). Its rostrolateral process terminates in a bifurcated facet for articulation with the postorbital."
http://icb.oxfordjournals.org/cgi/content/full/40/4/687
"The most important bird from the Liaoning deposits in China is Confuciusornis the only bird besides Archaeopteryx with enlarged claws for climbing. It also has a full set of gastralia and a postorbital bone that meets the jugal." (
http://www.freethought-forum.com/forum/showthread.php?t=11592&garpg=3
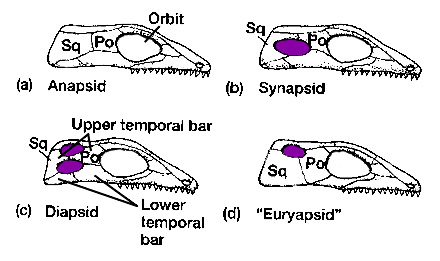
"In the drawing, the “orbit” is the eye socket, “Sq” indicates the squamosal bone, one of the bones that makes up the skull, and “Po” indicates the postorbital bone."
Summary
Time for a summary.
Here are the characteristics I have reviewed so far:
Here are the characteristics I have reviewed so far:
- Leg (including tibia/tibiotarsus, metatarsus/tarsometatarsus and femur)
- Post hatch care
- Pubic bones
- Wing composition (inluding actinofibrils, pycnofibres, fibrils, trailing "feathers", and ankles)
- Metabolic rate
- Beaks
- Crests
- Sexual maturation
- Trabeculae
- Wing bones (including carpus)
- Dentition
- Feet
- Mode of flight
- Furcula
- Acetabulum
- Warm blooded
- Keeled breastbone
- Synsacrum
- Notarium
- Pneumatization
- Uncinate process
- Gizzard stones
- Fused pelvic bones
- Vision
- Brain
- Landing
- Breathing
- Genome evidence
- Harry Seeley
- Lineage
- Timing
- Bird/pterosaur correspondences (including flamingo etc)
Monday, May 24, 2010
* Dinosaur legs are not similar
Notice that in almost every respect, dinosaur legs are different than modern bird legs.
http://www.innerbird.com/pelvic_girdle/pelvic_girdle.html
"The change in musculature has been accompanied by equally significant changes to the bird’s [dinosaur's] leg. Dinosaurs have {1}two-unit limbs in the sense that {2} two long bones of roughly equal length form the leg above the foot. The weight of the body is borne by the toes and the {3} independent metatarsals of the foot are {4}relatively short.
A similar kind of leg appears in Archaeopteryx and Confuciusornis but in more advanced [modern] birds, {1}the femur is shorter than the tibia and {2}fused metatarsals form a new segment that is similar in length to the tibia. {3}Both the tibia and the metatarsals are fused with neighbouring bones from the joints and been renamed tibiotarsus and tarsometatarsus to reflect their new structure. In the bird’s {4}three-unit leg, the femur is held in a roughly horizontal position so that it functions more like a forward extension of the hip bones and contributes little to the length of the bird’s stride. The new arrangement is very useful when the after part of the abdomen is swollen by the presence of a very large egg. (see gallery of eggs and hips)."
http://www.innerbird.com/pelvic_girdle/pelvic_girdle.html
"The change in musculature has been accompanied by equally significant changes to the bird’s [dinosaur's] leg. Dinosaurs have {1}two-unit limbs in the sense that {2} two long bones of roughly equal length form the leg above the foot. The weight of the body is borne by the toes and the {3} independent metatarsals of the foot are {4}relatively short.
A similar kind of leg appears in Archaeopteryx and Confuciusornis but in more advanced [modern] birds, {1}the femur is shorter than the tibia and {2}fused metatarsals form a new segment that is similar in length to the tibia. {3}Both the tibia and the metatarsals are fused with neighbouring bones from the joints and been renamed tibiotarsus and tarsometatarsus to reflect their new structure. In the bird’s {4}three-unit leg, the femur is held in a roughly horizontal position so that it functions more like a forward extension of the hip bones and contributes little to the length of the bird’s stride. The new arrangement is very useful when the after part of the abdomen is swollen by the presence of a very large egg. (see gallery of eggs and hips)."
Leg Similarities (4)
More on the pterosaur tibiotarsus:
http://www.springerlink.com/content/q977815700x18614/
http://www.springerlink.com/content/q977815700x18614/
Avian-like tibiotarsi of pterodactyloids (Reptilia: Pterosauria) from the Upper Jurassic of East Africa"Tibiotarsi of Dsungaripterus brancai (Reck) (Upper Jurassic, East Africa),D. weii Young (Lower Cretaceous, China) and Puntanipterus globosus Bonaparte & Sanchez (Lower Cretaceous, South America) have a bird-like distal end with attachment areas for a transverse ligament anteriorly, lateral and medial ligamentous prominences, and an anteroposteriorly, expanded pulley-like articular surface. The M. extensor digitorum longus flexed the ankle and probably also extended the digits as in living birds and mammals. A separate tendinous slip for digit I probably passed from the M. flexor digitorum longus in a groove posteroventral to the medial ligamentous prominence. " (Peter M. Galton)http://books.google.ca/books?id=idta6AVV-tIC&pg=PA11&lpg=PA11&dq=bird+metatarsus+pterosaur+metatarsal&source=bl&ots=2E_X9V4bPt&sig=0bLfNMhd8yejNSOafECEnBqKAyo&hl=en&ei=uOb6S_3OGsWclgeZ2oHdCg&sa=X&oi=book_result&ct=result&resnum=3&ved=0CB4Q6AEwAg#v=onepage&q&f=false
"The metamorphosis of the ankle joints of pterosaurs during development is interesting. In juvenile specimens, the proximal row consists of astragalus and calcaneum which are both reduced in length proximo-distally to become essentially only caps over the lower ends of the tibia, with the loss of the calcaneal tuber.http://pterosaur.net/myths.php
In adult [pterosaur] specimens, these two bones [the calcaneum and the astragalus] are fused with the tibia to form a rolling hinge joint at their distal condyles as seen in modern birds." (Sankar Chatterjee, R. J. Templin)
The shared presence of a relatively long neck with proportionally long cervical vertebrae, of an elongate tibia, and of a hinge-like ankle joint and elongate metatarsals in the foot indicate that pterosaurs, dinosaurs and a few other groups should be united in a clade which has been termed Ornithodira. Also see these references: http://pterosaurnet.blogspot.ca/2010/05/bird-and-pterosaur-ankles.html http://pterosaurnet.blogspot.ca/2010/05/more-on-ankles.html |
Leg Similarities (3)
The modern bird tarsometatarsus.
There are two kinds of fusion involved. The metatarsal bones themselves fuse together and they in turn fuse with some small anklebones (tarsals) forming the tarsometatarsus.
http://geolmag.geoscienceworld.org/cgi/content/abstract/139/4/421
http://www.innerbird.com/pelvic_girdle/pelvic_girdle.html#avian_leg
-->
http://geolmag.geoscienceworld.org/cgi/content/abstract/139/4/421
"There are several ways and extents in which the tarsometatarsal fusion is accomplished throughout bird evolution. Namely, in Neornithes (modern birds) the fusion is most thorough at the distal (metatarsal) end though present along the entire length of the bone. In the Enantiornithes, a group of Mesozoic birds, the fusion was complete at the proximal (tarsal) end but the distal metatarsi still were somewhat separate."
http://www.innerbird.com/pelvic_girdle/pelvic_girdle.html#avian_leg
"The third long bone in the [modern bird] leg is the tarsometatarsus. It is unique to birds and consists of the fused remnants of the metatarsal bones (long bones of the foot) and some small anklebones or tarsals. It is thought [incorrectly] to have evolved from comparable bones in dinosaurs that also walked on their toes. In many birds, the tarsometatarsus is the only bone that extends beyond the skirt of feathers. Like other parts of the avian foot, it has no muscles of its own but includes grooves and other structures to guide long tendons that stretch between the toes and the thigh muscles."
-->
Leg Similarities (2)
Tibiotarsus:
http://www.innerbird.com/pelvic_girdle/pelvic_girdle.html#avian_leg
"Birds are the only living vertebrate whose hind limb includes three long bones in sequence. The innermost of the two long bones are similar to those found in most vertebrates. At the hip, the femur, is held more or less parallel to the ground and is bound to the hipbones by the massive thigh muscles. In effect, the femur is an addition to the hipbones and its rotation contributes little to the length of the bird’s stride. The second long bone in the leg is the tibiotarsus. As the name implies, it is the fused product of the tibia and some tarsal bones. In other vertebrates, the tibia lies parallel to the fibula, another long bone of similar size. The two bones act together to facilitate rotation at the ankle. In birds there is no rotation at the ankle and the fibula no longer reaches the ankle joint and no longer qualifies as a major long bone."
Here is an additional reference:
http://sp.lyellcollection.org/cgi/content/abstract/217/1/23
"The tarsus of Triassic pterosaurs consists of two proximal tarsals, which fuse to the tibia during ontogeny, forming a tibiotarsus, and two distal tarsals." (Fabio M. Dalla Vecchia)
Also see the post entitled Leg Similarities (1) demonstrating that the pterosaur and modern bird tibiotarsus are near identical in terms of the tibia and the fibula.
* Post-hatching care
We saw in the earlier post that:
"The single most astounding feature of dinosaur pubic bones is that they formed a solid ring with a rather small opening."
Here is the expanded description of this situation, which shows another difference between dinosaurs and modern birds:
http://www.innerbird.com/pelvic_girdle/pelvic_girdle.html#pubic_bone
"It is possible to re-interpret the structural changes in the avian hip and tail as changes related to the evolutionary benefits offered by increased parental investment in offspring. The single most astounding feature of dinosaur pubic bones is that they formed a solid ring with a rather small opening. That opening severely limited the size of the egg and, therefore, the size of the hatchling. This anatomical bottleneck limited dinosaurs to reproductive strategies involving relatively large numbers of small young. Hatchlings may have needed foods and habitats that were different from those of the adult animals. Most animals that lay large clutches of eggs have post-hatching care by the adult that is somewhat hit-and-miss as it is in modern crocodiles, and not at all like the intense, dedicated effort made by many modern birds."
The fused pubic bones of the dinosaur led to dinosaur "post-hatching care" that was "not at all like many modern birds".
"The single most astounding feature of dinosaur pubic bones is that they formed a solid ring with a rather small opening."
Here is the expanded description of this situation, which shows another difference between dinosaurs and modern birds:
http://www.innerbird.com/pelvic_girdle/pelvic_girdle.html#pubic_bone
"It is possible to re-interpret the structural changes in the avian hip and tail as changes related to the evolutionary benefits offered by increased parental investment in offspring. The single most astounding feature of dinosaur pubic bones is that they formed a solid ring with a rather small opening. That opening severely limited the size of the egg and, therefore, the size of the hatchling. This anatomical bottleneck limited dinosaurs to reproductive strategies involving relatively large numbers of small young. Hatchlings may have needed foods and habitats that were different from those of the adult animals. Most animals that lay large clutches of eggs have post-hatching care by the adult that is somewhat hit-and-miss as it is in modern crocodiles, and not at all like the intense, dedicated effort made by many modern birds."
The fused pubic bones of the dinosaur led to dinosaur "post-hatching care" that was "not at all like many modern birds".
Subscribe to:
Comments (Atom)