The cladogram shows that birds did not evolve from dinosaurs.
Click to enlarge.

See Comments for taxa and characters.
The cladogram shows that birds did not evolve from dinosaurs.
Click to enlarge.
Various studies analyze dinosaurs (including birds that they think evolved from dinosaurs).
And other studies analyze pterosaurs.
But no studies include both dinosaurs and pterosaurs scored on the same characteristics.
The one exception is the 2022 study which includes the analysis of one character - feather type, for both pterosaurs and dinosaurs.
https://zenodo.org/record/6122213#.YqC5YP_MKcx
See tab 4 of spreadsheet.
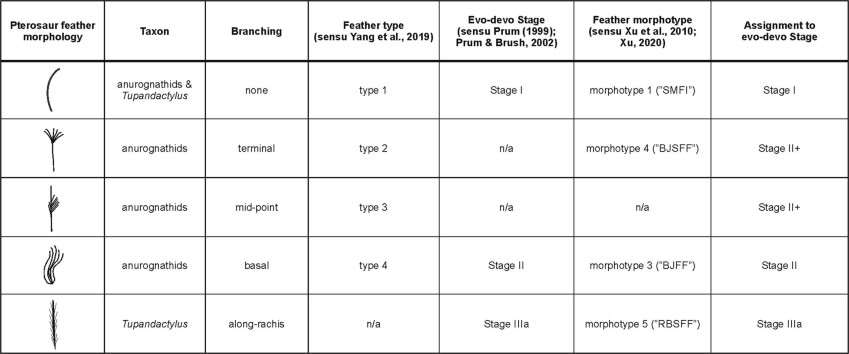
Notice that Tupandactylus is scored as feather type 5.

Consider the short tailed pterosaurs (eg. tupandactylus) and the short tailed scansoriopterygids (eg epidexipteryx).
Pterosaurs like birds, were capable of powered flight. It seems that command of the skies is not the only thing that these two types of vertebrate had in common. Thanks to a remarkable series of discoveries from the remote Turpan-Hami Basin located in the Xinjiang Uygur Autonomous Region (north-western China), palaeontologists have learned that Pterosaurs, like many living birds nested in colonies, that they had preferred nesting sites and when young, Pterosaurs needed a degree of parental care, just like many species of birds today.
Significantly, the number of eggs discovered are far too many to have been laid by a single female. This suggests that these flying reptiles nested in colonies and furthermore, the overlaying of multiple clutches of eggs indicates that Pterosaurs, like many birds today, returned to the same nesting sites each year. As the authors conclude, “the similarity between these groups goes beyond wings”.
The study:
https://www.science.org/doi/10.1126/science.aan2329
The scientific paper: “Egg Accumulation with 3D Embryos Provides Insight into the Life History of a Pterosaur”
Here is a summary of the basic ideas of this site:
Birds developed in a lineage from short bony tailed pterosaurs to short bony tailed scansoriopterygidae that developed into basal pygostylia birds that developed into modern birds. A side lineage earlier developed from the long tailed pterosaurs to long bony tailed scansoriopterygidae. That linage went extinct.
https://biblio.ugent.be/publication/8752596/file/8752597.pdf
Some feather morphologies are shared (that is, monofilaments, brush-like and tufted feathers and feathers with along-rachis branching), but others are not—for example, feathers with midpoint branching in pterosaurs and all feathers with barbules in theropods. Barbules are thus a unique innovation of theropod feathers.
This would be more accurate to say:
all feathers with barbules in
theropods[Paraves]. Barbules are thus a unique innovation oftheropod[Paraves] feathers.
Paraves* did not develop from any dinosaurs.
" including secondarily flightless oviraptorids and secondarily flightless ornithomimosaurs
Similar skull openings of pterodactylid and scansoriopterygid:

https://en.wikipedia.org/wiki/File:Epidexipteryx.jpg

High level similarities:
Pterosaur pterodactylid (eg tupandactylus):
Membrane wing
Longest 4th finger
Stage IIIa feathers
Short bony tail
Paraves scansoriopterygid (eg epidexipteryx):
Membrane wing
Longest 4th finger
Stage IIIa feathers
Short bony tail
Significant evidence that short bony tailed pterodactylid developed into short bony tailed scansoriopterygid.
https://biblio.ugent.be/publication/8752596/file/8752597.pdf
To our knowledge, stage IIIa feathers have not previously been reported in pterosaurs. The Tupandactylus branched structures resemble those in the dromaeosaurid dinosaur Sinornithosaurus millenii27, which are considered homologous to avian feathers28, and differ from the three types of branched feathers described in anurognathid pterosaurs2 .
This mode of branching is directly comparable to that in stage IIIa feathers19,20 of extant birds, that is, with barbs branching from a central rachis. This is strong evidence that the fossil branched structures are feathers comprising a rachis and barbs.
In other words the pterosaur Tupandactylus stage IIIa feathers resembled those in Sinornithosaurus (paraves/dromaeosauridae) which are considered homologous to avian feathers.
This is exceptional support for the pterosaur to bird theory.

https://biblio.ugent.be/publication/8752596/file/8752597.pdf (2022)
The genotypic and phenotypic characters could both be ancestral to avemetatarsalians; alternatively, both evolved independently in theropods and pterosaurs, or the genes are ancestral and the phenotypic expression occurred independently in the two groups. Our ancestral-state estimations (Extended Data Fig. 9e) reveal that the most parsimonious scenario is that feathers in the avemetatarsalian ancestor had melanosomes with different geometries. This is consistent with a single, deep evolutionary origin for this feature, whereby critical shifts in the genetic machinery facilitating plasticity in melanosome shape occurred in the common ancestor of pterosaurs and birds. Key genomic controls on melanin-based colouration that define the plumage colours of theropods and fossil and extant birds were therefore already in place in early-diverging avemetatarsalians in the Middle to Late Triassic.
Data availability
Additional data on melanosome geometry and the character matrix used in the phylogenetic analyses are available in the Zenodo.org data repository at https://doi.org/10.5281/zenodo.6122213. SEM images and samples are available from the corresponding authors on request.
https://biblio.ugent.be/publication/8752596/file/8752597.pdf (2022)
Remarkably well-preserved soft tissues in Mesozoic fossils have yielded substantial insights into the evolution of feathers(1). New evidence of branched feathers in pterosaurs suggests that feathers originated in the avemetatarsalian ancestor of pterosaurs and dinosaurs in the Early Triassic(2), but the homology of these pterosaur structures with feathers is controversial(3,4). Reports of pterosaur feathers with homogeneous ovoid melanosome geometries(2,5) suggest that they exhibited limited variation in colour, supporting hypotheses that early feathers functioned primarily in thermoregulation(6). Here we report the presence of diverse melanosome geometries in the skin and simple and branched feathers of a tapejarid pterosaur from the Early Cretaceous found in Brazil. The melanosomes form distinct populations in different feather types and the skin, a feature previously known only in theropod dinosaurs, including birds. These tissue-specific melanosome geometries in pterosaurs indicate that manipulation of feather colour-and thus functions of feathers in visual communication-has deep evolutionary origins. These features show that genetic regulation of melanosome chemistry and shape(7-9) was active early in feather evolution.

Developmental Model of the Origin and Diversification of Feathers
A predicted transition series of feather follicles based on the hypothesized series of evolutionary novelties in feather developmental mechanisms (Figure 4) from Prum (1999). Stage I—Origin of an undifferentiated tubular collar yields the first feather, a hollow cylinder. Stage II—Origin of a collar with differentiated barb ridges results in a mature feather with a tuft of unbranched barbs and a basal calamus emerging from a superficial sheath. Stage IIIa—Origin of helical displacement of barb ridges and the new barb locus results in a pinnate feather with an indeterminate number of unbranched barbs fused to a central rachis. Stage IIIb—Origin of peripheral barbule plates within barb ridges yields a feather with numerous branched barbs attached to a basal calamus. Stages IIIaIIIb—Origin of a feather with both a rachis and barbs with barbules creates a bipinnate, open pennaceous structure.
Morphotype 5 is only known in Epidexipteryx. It consists of parallel barbs arising from the edge of a membrane structure (Zhang et al., 2008b).
Video from Maria McNamara:
https://www.youtube.com/watch?v=Bmm3IyyjcS0
Difference between feather (with no barbules) and down:
https://www.youtube.com/watch?v=5rcwajqH0Dc&t=1s
http://link.springer.com/article/10.1007/s10336-014-1098-9/fulltext.html (2014)
Jurassic archosaur is a non-dinosaurian bird
Stephen A. Czerkas, Alan Feduccia
Re-examination utilizing Keyence 3D digital microscopy and low angled illumination of the fossil Scansoriopteryx, a problematic sparrow-size pre-Archaeopteryx specimen from the Jurassic Daohugou Biotas, provides new evidence which challenges the widely accepted hypothesis that birds are derived from dinosaurs in which avian flight originated from cursorial forms. Contrary to previous interpretations in which Scansoriopteryx was considered to be a coelurosaurian theropod dinosaur, the absence of fundamental dinosaurian characteristics demonstrates that it was not derived from a dinosaurian ancestry and should not be considered as a theropod dinosaur.
I am suggesting the following:
Long bony tailed rhamphorhynchid --> Long bony tailed scansoriopterygid (eg scansoriopteryx) --> Derived long bony tailed Paraves (eg Anchiornithids) --> Extinction
Long bony tailed rhamphorhynchid --> Short bony tailed pterodactylid --> Short bony tailed scansoriopterygid (eg epidexipteryx) --> Pygostylia --> Modern birds.
To this point I have focussed on the development from long bony tailed pterosaurs (rhamphorhynchids) to long bony tailed scansoriopterygids (eg scansoriopteryx).
I have not followed on from there to whether long bony tailed Paraves became short bony tailed pygostylia. I suggest they did not.
And I have not talked about the short tailed pterodactylids.
I suggest they are related. Short tailed pterosaurs (eg pterodactylids) developed into short bony tailed scansoriopterygids (eg epidexipteryx). Which then developed into pygostylia.
Nature experimented with various forms of flight. A big challenge was transitioning from long bony tail to short bony tail. This was accomplished WITHIN pterosaur lineage.
https://en.wikipedia.org/wiki/Pterodactyloidea
"Pterodactyloidea is traditionally considered to be the group of short-tailed pterosaurs with long wrists (metacarpus), compared with the relatively long tails and short wrist bones of basal pterosaurs ("rhamphorhynchoids")."
https://en.wikipedia.org/wiki/Scansoriopterygidae
"Epidexipteryx had a short tail (70% the length of the torso), anchoring long tail feathers, while Scansoriopteryx had a very long tail (over three times as long as the torso) with a short spray of feathers at the tip."
Some long bony tailed pterosaurs (eg rhamphorhynchids) developed into long bony tailed scansoriopterygids. And some of those long bony tailed scansoriopterygids developed into other more derived long bony tailed members of Paraves. Then they went extinct.
But there are two paths. One from long bony tailed pterosaurs and the other from (later) short bony tailed pterosaurs (eg pterodactylids)
Some short bony tailed pterosaurs (eg pterodactylids) developed into short bony tailed scansoriopterygids. And some of the short bony tailed scansoriopterygids developed into pygostylia.
This means that later stage feathers (after stage IIIa) developed in both lines.
This suggests there was greater experimentation with wing-assisted locomotion before theropod flight evolved than previously appreciated. This study adds invaluable support for multiple origins of powered flight potential in theropods (≥3 times), which we now know was from ancestors already nearing associated thresholds, and provides a framework for its further study
{kind=link}
{kind=link}
{kind=link}