Here is a reference that shows the internal composition of the pterosaur wing membrane. It is quite astonishing. Check out page 241 and the pages nearby.
http://books.google.ca/books?id=8CKYxcylOycC&pg=PA243&lpg=PA243&dq=tenopatagium&source=bl&ots=SopV9CAGec&sig=-gWOltWiFGplrU9tcXV8X8pBPUI&hl=en&ei=vyTjS5aEBoT6lwf_uMC9Ag&sa=X&oi=book_result&ct=result&resnum=6&ved=0CC8Q6AEwBQ#v=onepage&q=tenopatagium&f=false
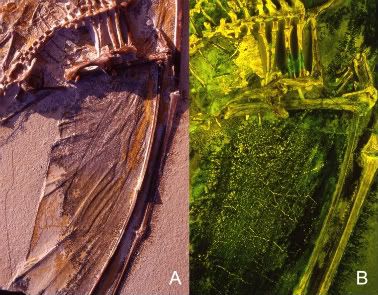
Page 243:
The strings cross the fibres at angles between 30° and 90°
http://archosaurmusings.files.wordpress.com/2008/07/darkwing.jpg?w=764&h=510
It is astonishing, because we can see that the structure and material of feathers is already present as the actinofibrils in the membrane.
http://www.ucmp.berkeley.edu/vertebrates/flight/pter.html
"Bird feathers are analogous to the wing fibers of pterosaurs,"
PTEROSAURS
http://www.springerlink.com/content/v8434087565413kk/fulltext.pdf?page=1
"Wellnhofer [4, 5] and Padian [6, 7], following von Zittel [8],described a system of fine structural fibers investing the wing membrane, in a pattern similar to the orientation of the feather shafts of birds and the wing fingers of bats, both principal structural elements supporting the patagium and responsible for the transmission of aerodynamic force."
http://www.jstor.org/pss/2400656
The wing membrane was supported and controlled through a system of stiffened, intercalated fibers, which were oriented like the main structural elements in the wings of birds and bats.
From David Unwin's book "The Pterosaurs from Deep Time":
https://archive.org/stream/The_Pterosaurs_From_Deep_Time_by_David_M._Unwin/The_Pterosaurs_From_Deep_Time_by_David_M._Unwin_djvu.txt
"...[T]he wing fibers were embedded within the patagia [wing membranes] and typically measured a little less than one-tenth of a millimetre in diameter- about twice the thickness of a human hair. In some spots unravelled fibers reveal that they were composite structures composed of at least 20 or 30 very fine strands, wound together in a helical fashion. Each strand was only a few hundredth of a millimeter across and probably made of collagen a material that is common in the skin of vertebrates".
http://en.wikipedia.org/wiki/Pterosaur
"The actual function of the actinofibrils is unknown, as is the exact material from which they were made. Depending on their exact composition (keratin, muscle, elastic structures, etc.), they may have been stiffening or strengthening agents in the outer part of the wing.[6] The wing membranes also contained a thin layer of muscle, fibrous tissue, and a unique, complex circulatory system of looping blood vessels.[7]"
"Since they [actinofibrils] were external, they were probably epidermal structures composed of keratin as in scales and feathers."
http://en.wikipedia.org/wiki/Pterosaur
"research has since shown that the wing membranes of pterosaurs were actually highly complex and dynamic structures suited to an active style of flight. First, the outer wings (from the wing to the elbow) were strengthened by closely spaced fibers called actinofibrils.[5] The actinofibrils themselves consisted of three distinct layers in the wing, forming a crisscross pattern when superimposed on one another."
http://en.wikipedia.org/wiki/Pterosaur
"The brachiopatagium ("arm membrane") was the primary component of the [pterosaur] wing, stretching from the highly elongated fourth finger of the hand to the hind limbs"
BIRDS
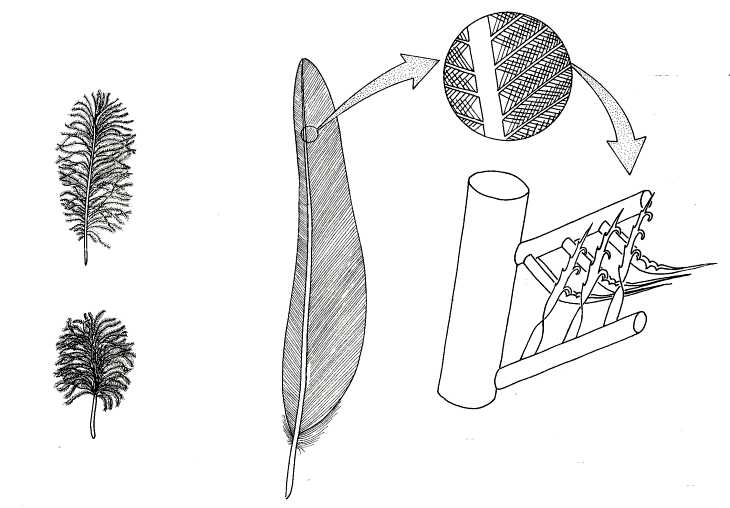
http://en.wikipedia.org/wiki/Pennaceous_feather
"Pennaceous feathers are also known as contour feathers. This type of feather is present in most modern birds and has been shown in some species of maniraptoran dinosaurs. A pennaceous feather has a stalk or quill. Its basal part, called a calamus, is embedded in the skin. The calamus is hollow and has pith formed from the dry remains of the feather pulp, and the calamus opens below by an inferior umbilicus and above by a superior umbilicus. The stalk above the calamus is a solid rachis having an umbilical groove on its underside. Pennaceous feathers have a central shaft (or rachis) with vanes or vaxillum spreading to either side. These vanes are composed of a high number of flattened barbs, that are connected to one another with barbules.
The barbules are tiny strands that criss-cross on the flattened sides of the barbs. This forms a kind of miniature velcro-like mesh that holds all the barbs together, stabilizing the vanes."
http://en.wikipedia.org/wiki/Flight_feather
Remiges (from the Latin for "oarsman") are located on the posterior side of the [bird] wing. Ligaments attach the long calami, or quills, firmly to the wing bones, and a thick, strong band of tendinous tissue—known as the postpatagium—helps to hold and support the remiges in place.[1]
http://www.wordnik.com/words/postpatagium
" In ornithology, the triangular fold of skin, just back of the shoulder-joint, which runs from the side of the body to the upper posterior face of the upper arm."
http://www.innvista.com/science/zoology/ornithology/birdtopo.htm
"Postpatagium is the tough band of tendinous tissue that envelops and supports the quills of all the wing remiges, from elbow to wingtip. The postpatagium provides much of the elastic strength of the wing and keeps the flight feathers properly aligned and firmly attached to the wing skeleton."
This is interesting:
http://dml.cmnh.org/2001Jan/msg00135.html
"I`m not sure exactly how feathers developed,but actinofibrils arranged in a pattern similar to flight feather rachis is very suggestive."
Earlier post:
http://pterosaurnet.blogspot.ca/2010/05/feathers.html
FOR REFERENCE:
http://www.biology-resources.com/drawing-bird-feather-structure.html
http://www.glogster.com/media/5/34/29/19/34291999.jpg
http://www.ucmp.berkeley.edu/vertebrates/flight/pterosaurwing.gif
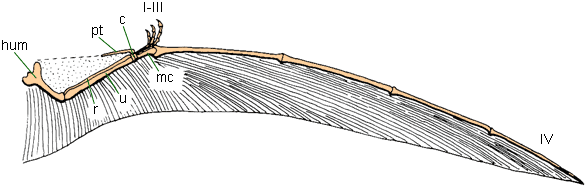
http://upload.wikimedia.org/wikipedia/commons/thumb/d/d3/Pterosaur_wing_BW2.jpg/300px-Pterosaur_wing_BW2.jpg

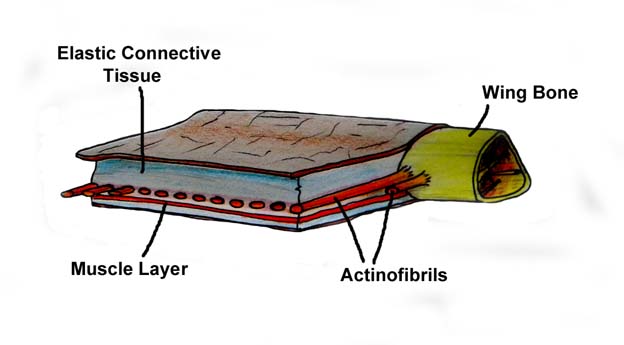














{kind=link}
{kind=link}