BASAL PARAVES
http://www.pnas.org/content/107/7/2972.full.pdf+html
The absence of an antitrochanter and a supra acetabular shelf (SAC) in the eumaniraptorans, including dromaeosaurids,would make elevation and splaying of the legs even easier (8).
The [Archaeopteryx] pelvis has an incompletely open acetabulum, and there is no characteristic dinosaurian supra-acetabular shelf. The femoral head turns forwards rather than extending perpendicular to the shaft.(Martin)
Medially, the medial shelf of the brevis fossa is split into separate anterior and posterior processes, a condition unique to Hesperonychus. The acetabulum is similar to those of other dromaeosaurids in that it lacks a prominent supracetabular crest (30, 36).
Unlike theropod dinosaurs, invariably exhibiting acompletely perforated and open acetabulum, Scansoriopteryxhas a partially closed acetabulum, and no sign of a
supra-acetabular shelf or an antitrochanter.
PTEROSAUR
The hip joint of pterosaurs is more mobile and profoundly different from that of theropods but is reminiscent of that of mammals, especially of humans, allowing a wide range of adduction and abduction in the vertical plane
The femoral component [of the Anhanguera pterosaur femur] is a well defined spherical head which is distinctly separated from the shaft by a narrow non-articular neck at an obtuse angle of 160 degrees.
The head forms a ball and socket joint with the close-fitting, shallow and imperforate acetabulum.
In this orientation, there is no overhanging acetabular rim to support the femur in a vertical position.
pterosaurs had no antitrochanter locking device to keep the femur in a near-horizontal position.
DINOSAUR
http://blogs.scientificamerican.com/WSS/post.php?blog=33&post=10113
In the theropod [dinosaur] there are proximally placed trochanters on the femur, and supra-acetabular shelves and antritrochanters on the ilium
MODERN BIRD (for reference)
http://connection.ebscohost.com/c/articles/26133921/antitrochanter-birds-form-function-balance
The antitrochanter serves as a brace to prevent abduction of the hindlimb and to absorb stresses that would otherwise be placed on the head of the femur during bipedal locomotion. The drum-in-trough-like form of the antitrochanter-femur articulation tends to assist in the transfer of long-axis rotational movements of the femur to the pelvis. The avian antitrochanter is a derived feature of birds that evolved as an aid in maintaining balance during bipedal terrestrial locomotion.
ACETABULUM
Dinosaurs
http://www.ucmp.berkeley.edu/diapsids/dinomm.html
"One important dinosaurian synapomorphy is the perforate [completely open] acetabulum, simply a "hip bone" (actually three connected bones, together called the pelvis) with a hole in the center where the head of the femur ("thigh bone") sits. This construction of the hip joint makes an erect stance (hindlimbs located directly beneath the body) necessary — like most mammals, but unlike other reptiles which have a less erect and more sprawling posture. Dinosaurs are unique among all tetrapods in having this [completely] perforate acetabulum."http://en.wikipedia.org/wiki/Acetabulum
The acetabulum is a concave surface of the pelvis. The head of the femur meets with the pelvis at the acetabulum, forming the hip joint.
In reptiles and in birds, the acetabula are deep sockets. Organisms in the dinosauria clade are defined by a perforate [completely open] acetabulum, a hip socket with a hole at the center of its inside surface.[1]"
http://www.aou.org/auk/content/130/1/0001-0013.pdf
A partially closed acetabulum is seen in basal archosaurs andhttp://dml.cmnh.org/2005May/msg00317.html
is characteristic of the scansoriopterids and Jurassic feathered
forms such as Anchiornis initially described as near Aves by Xu et al. (2009).
This results in a somewhat sprawling position for the [Archaeopteryx] femur that is corrected at the knee joint, resulting in a functionally vertical leg. (Martin)
The pelvis has an incompletely open acetabulum, and there is no characteristic dinosaurian supra-acetabular shelf. The femoral head turns forwards rather than extending perpendicular to the shaft. (Martin)https://answers.yahoo.com/question/index?qid=20131108220005AAf5Wev
In his latest book, "Riddle of the Feathered Dragons", Feduccia (p.160) points out that Anchiornis does not have a fully open acetabulum, which makes it different than the fully open acetabulum of dinosaurs. Unlike theropods, it does not have a supra-acetabular shelf even though the authors who described it claim that it does. Feduccia says it looks more like a slight rim and is certainly not like any typical theropods. If anything, these fossils exhibit clear differences with dinosaurs in those traits that are typically found in dinosaurs.
http://blogs.scientificamerican.com/tetrapod-zoology/2013/11/18/flight-of-the-microraptor/
(Holtz comment)
You can infer humans can do splits because we, like many mammals, have a ball-and-socket joint between the femoral head and the acetabulum. Dinosaurs (including Microraptor and birds) have a different form of articulation: more of a roller-joint than a ball-and-socket.Also sprawling is covered on page 183 and page 251 of Feduccia's book "Riddle of the Feathered Dragons".
http://www.pnas.org/content/106/13/5002.full
The medial opening of the acetabulum is partially closed, as it is in other Dromaeosauridae (36). The acetabulum opens dorsolaterally rather than laterally, as is the case in Velociraptor (38), suggesting the ability to partially abduct the hindlimbs. This morphology is of interest in light of proposals that Microraptor gui abducted its feathered hindlimbs to function as airfoils (24).PTEROSAUR etc
http://www.jstor.org/pss/4523298
As Unwin (1988) pointed out, pterosaurs have an imperforate acetabulum,
Page 83.
http://bigcat.fhsu.edu/biology/cbennett/flotsam/Bennett-1990-Santana-pelvis.pdf
http://books.google.ca/books?id=idta6AVV-tIC&pg=PA10&lpg=PA10&dq=Unwin+%281988%29+,+pterosaurs+have+an+imperforate+acetabulum,&source=bl&ots=2E_W7T0hQs&sig=sEqzrX-ZuACLWeCHl23YaV9z9gc&hl=en&ei=2TvoS6W2B4bGlQfmxfnXAw&sa=X&oi=book_result&ct=result&resnum=3&ved=0CCAQ6AEwAg#v=onepage&q=Unwin%20%281988%29%20%2C%20pterosaurs%20have%20an%20imperforate%20acetabulum%2C&f=false
See page 10.
THIS DESCRIBES THE DIFFERENCE BETWEEN DINOSAUR AND PTEROSAUR FEMUR ARTICULATION
Note that the dinosaur joint is not ball and socket, as it is in pterosaurs and birds.
"In theropods, the femoral component is cylindrical without any distinctive head and neck."
"The hip joint of pterosaurs is more mobile and profoundly different from that of theropods but is reminiscent of that of mammals, especially of humans, allowing a wide range of adduction and abduction in the vertical plane"
"The femoral component [of the Anhanguera pterosaur femur] is a well defined spherical head which is distinctly separated from the shaft by a narrow non-articular neck at an obtuse angle of 160 degrees.
The head forms a ball and socket joint with the close-fitting, shallow and imperforate acetabulum."
Also, there seems to be a difference in the orientation of the pelvis of rhamphorhynchoids and pterodactyloids.
See page 84 of this reference:
http://bigcat.fhsu.edu/biology/cbennett/flotsam/Bennett-1990-Santana-pelvis.pdf
http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0058451

The specimen [Vectidraco daisymorrisae] includes most of the left side of the articulated pelvis (Figure 1): the prepubis, ventral part of the pubis and part of the anterior process of the ilium are missing, and adhering matrix (which cannot be removed without damaging the specimen) obscures the interior of the acetabulum and much of the medial surface of the pelvis.
http://pterosaur.net/fossils/coloborhynchus_pelvis_2.jpg
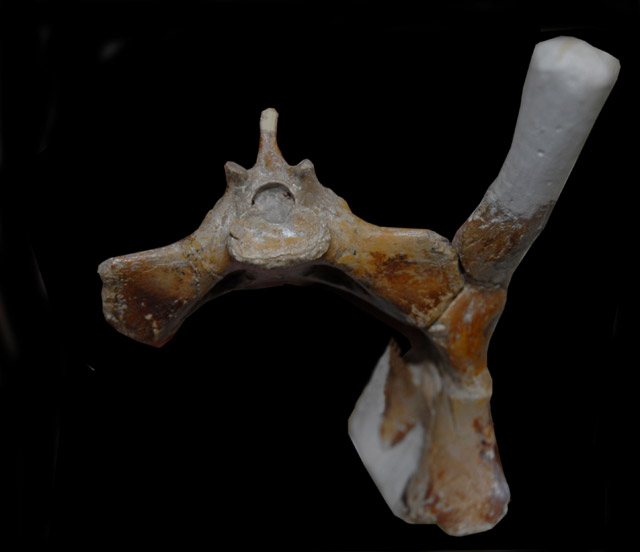
http://pterosaur.net/fossils/coloborhynchus_pelvis.jpg

MODERN BIRD
See page 30 of this reference:
http://books.google.ca/books?id=YhAY...mammal&f=false
Quote:
| The right angled ball-and-socket joint that forms the [bird] hip |
FOR REFERENCE:
http://en.wikipedia.org/wiki/Hip
http://en.wikipedia.org/wiki/Femur

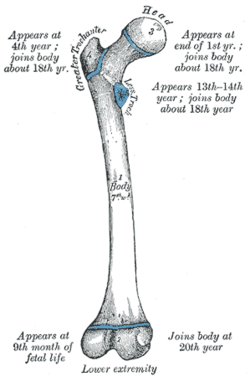
Dinosaur

Chicken femur:

http://www.pnas.org/content/104/5/1576.full
Microraptor gui, a four-winged dromaeosaur from the Early Cretaceous of China, provides strong evidence for an arboreal-gliding origin of avian flight. It possessed asymmetric flight feathers not only on the manus but also on the pes. A previously published reconstruction shows that the hindwing of Microraptor supported by a laterally extended leg would have formed a second pair of wings in tetrapteryx fashion. However, this wing design conflicts with known theropod limb joints that entail a parasagittal posture of the hindlimb.
The sources I have looked at say that birds have a perforated acetabulum.
ReplyDeleteSee:
http://books.google.com/books?id=yS_dj0P6XzwC&pg=PA77&lpg=PA77&dq=perforated+acetabulum+bird&source=bl&ots=8kRq6ggk18&sig=ulfF3CTVAbLjC16VH3lw8nJpQgo&hl=en&sa=X&ei=ts8GUbutDca60AH6w4DICA&ved=0CFQQ6AEwBw#v=onepage&q=perforated%20acetabulum%20bird&f=false
And these much older sources:
http://books.google.com/books?id=6xYCAAAAYAAJ&pg=PA510&lpg=PA510&dq=perforated+acetabulum+bird&source=bl&ots=kRHID7nEcJ&sig=u5JDvhMsoqCZVyrIGR_0HpMCINQ&hl=en&sa=X&ei=dNEGUbu2NMqE0QG13IDwAw&ved=0CD4Q6AEwBDgK#v=onepage&q=perforated%20acetabulum%20bird&f=false
http://books.google.com/books?id=oG0NAQAAMAAJ&pg=PA98&lpg=PA98&dq=perforated+acetabulum+bird&source=bl&ots=KmExgGoeBE&sig=r0QyDiuzTr1vmijUE_yKdxp9sW4&hl=en&sa=X&ei=ts8GUbutDca60AH6w4DICA&ved=0CFEQ6AEwBg#v=onepage&q=perforated%20acetabulum%20bird&f=false
I have looked at the first reference. It indicates that dinosaurs had a perforated acetabulum as I mentioned. It does not indicate that birds have a perforated acetabulum.
ReplyDeleteDiogenes - please elaborate with a copy and paste, what leads you to the conclusion that birds have a perforated acetabulum.
Google books cannot be copied and pasted, so I will quote them as best I can.
ReplyDeletehttp://books.google.com/books?id=6xYCAAAAYAAJ&pg=PA510&lpg=PA510&dq=perforated+acetabulum+bird&source=bl&ots=kRHID7nEcJ&sig=u5JDvhMsoqCZVyrIGR_0HpMCINQ&hl=en&sa=X&ei=dNEGUbu2NMqE0QG13IDwAw&ved=0CD4Q6AEwBDgK#v=onepage&q=perforated%20acetabulum%20bird&f=false
Page 510, 2nd paragraph:
"The Pelvis of A Bird... the acetabulum is perforated by a wide foramen."
http://books.google.com/books?id=oG0NAQAAMAAJ&pg=PA98&lpg=PA98&dq=perforated+acetabulum+bird&source=bl&ots=KmExgGoeBE&sig=r0QyDiuzTr1vmijUE_yKdxp9sW4&hl=en&sa=X&ei=ts8GUbutDca60AH6w4DICA&ved=0CFEQ6AEwBg#v=onepage&q=perforated%20acetabulum%20bird&f=false
Page 98, 1st paragraph.
"...in Birds, the acetabulum is perforated."
What do they mean by "bird" in these quotes. Please give the specific taxa they are referring to.Then we can get more info on those specific taxa.
ReplyDeleteBy the way I notice that you are now referring to the second and third quotes, not the first one. Why not start with the first one as I did?
From the first link:
Deletehttp://books.google.com/books?id=yS_dj0P6XzwC&pg=PA77&lpg=PA77&dq=perforated+acetabulum+bird&source=bl&ots=8kRq6ggk18&sig=ulfF3CTVAbLjC16VH3lw8nJpQgo&hl=en&sa=X&ei=ts8GUbutDca60AH6w4DICA&ved=0CFQQ6AEwBw#v=onepage&q=perforated%20acetabulum%20bird&f=false
"Paleontologists have observed that all these characters defining dinosaurs are also present in birds. Characters such as the perforated acetabulum... support the position of Archaeopteryx within the dinosauria."
Yes.
ReplyDeleteI would really like to see for myself if the acetabulum is perforated or not. So the next time I cook a whole chicken or turkey, I will take photos. :)
I would be very interested in actual references (at least wikipedia) that answers these questions.
ReplyDeleteSo far the references I gave in the post seem to support what I am saying.
http://en.wikipedia.org/wiki/Acetabulum
ReplyDelete"In reptiles and in birds, the acetabula are deep sockets. Organisms in the dinosauria clade are defined by a perforate acetabulum, a hip socket with a hole at the center of its inside surface.[1]"
I hope Diogenes returns. This is an interesting discussion.
ReplyDeleteI have been unable to find the primary material of that Wikipedia quote. The source is cited, but the citation (the "[1]"), but the citation does not link to any reading material.
ReplyDeleteHere are some interesting illustrations.
One is a general illustration of bird hips, the other is specifically the rock pigeon hip.
http://etc.usf.edu/clipart/58400/58419/58419_pelvis.htm
http://etc.usf.edu/clipart/48600/48600/48600_pigeon_innom.htm
Perhaps the Wikipedia authors meant to link to this?
ReplyDeletehttp://www.ucmp.berkeley.edu/diapsids/dinomm.html
I am not sure.
Almost halfway down the page on the following website, there are two photographs of the hips of an emu (though if I recall correctly, you do not classify emu as birds).
http://bio.sunyorange.edu/updated2/comparative_anatomy/anat_3/l_hip.htm
I'll continue my bird hip photo hunt later.
It is an interesting discussion.
ReplyDeletehttp://en.allexperts.com/q/Paleontology-Dinosaurs-1571/modern-birds-still-possess.htm
The perforate acetabulum in dinosaurs is shared by primitive birds, including Archaeopteryx, but the feature is not present in modern birds. In fact the entire pelvis of birds has undergone extensive evolution since the days of Archaeopteryx, with the ilium, ischium and pubis having fused into one piece (the innominate bone). The hole which was once present closed up prior to the appearance of the group to which all modern birds belong (the Neornithes).
Similarly put:
Deletehttp://dml.cmnh.org/2006Oct/msg00042.html
"> From: nooj
> Subject: Do birds still have a perforate acetabulum?
> To: dinosaur@usc.edu
>
> Do modern birds still possess a perforate
> acetabulum? I wasn't able to find
> this information anywhere on the net. If so, can
> this be used as further
> evidence for the dinosaur-bird ancestry (which
> almost everyone agrees on
> anyway, but still).
Yes, modern birds do have a perforate acetabulum.
Eric Allen
UofIowa"
Still, both your quote and mine I find unsatisfying. I would like to find direct evidence either for or against the perforated acetabulum, such as a picture. Or an actual bird skeleton. It will probably be at least a couple weeks before I can get my hands on a whole chicken skeleton.
Similarly and somewhat humorously:
Deletehttp://dml.cmnh.org/2006Oct/msg00043.html
"From: owner-DINOSAUR@usc.edu [mailto:owner-DINOSAUR@usc.edu] On Behalf
Of nooj
Sent: Wednesday, October 04, 2006 11:56 PM
To: dinosaur@usc.edu
Subject: Do birds still have a perforate acetabulum?
Do modern birds still possess a perforate acetabulum? I wasn't able to
find this information anywhere on the net. If so, can this be used as
further evidence for the dinosaur-bird ancestry (which almost everyone
agrees on anyway, but still).
Thank you.
----- Original Message ----
From: "Ken.Carpenter@dmns.org"
To: soixmoi@gmail.com; dinosaur@usc.edu
Sent: Thursday, October 5, 2006 10:51:36 AM
Subject: RE: Do birds still have a perforate acetabulum?
Yes. Eat a chicken
Kenneth Carpenter, Ph.D.
Curator of Lower Vertebrate Paleontology/
Chief Preparator
Department of Earth Sciences
Denver Museum of Nature & Science
2001 Colorado Blvd.
Denver, CO 80205"
Emu are flightless birds.
ReplyDeleteI refer to them here:
http://pterosaurnet.blogspot.ca/2011/10/flightless-birds.html
Diogenes, it would be good to acquaint yourself with the material on the blog. That will save time and misunderstanding on your part.
One thing that I have not researched is whether the
ReplyDeleteRhamphorhynchoidea acetabulum was the same as or different than the Pterodactyloidea acetabulum