FLIGHTLESS BIRDS (Ratites)
- A Dromaeosaurid* subgroup --> Ornithomimosaurids --> Modern ratites (Ostrich (Struthio), Rhea (Rheidae), Cassowary, Emu (Casuariidae), Kiwi (Apteryx))
http://upload.wikimedia.org/wikipedia/commons/c/cd/Velociraptor_dinoguy2.jpg
Ornithomimid
http://upload.wikimedia.org/wikipedia/commons/thumb/3/39/Ornithomimid.jpg/800px-Ornithomimid.jpg
Ostrich
http://upload.wikimedia.org/wikipedia/commons/thumb/8/87/FlappingOstriches.jpg/643px-FlappingOstriches.jpg
Kiwi
http://0.tqn.com/d/goaustralia/1/0/9/i/kiwi.jpg
SEABIRDS (Ichthyornithes line)
- A Dromaeosaurid* subgroup -->
- An Enantiornithes seabird subgroup -->
- An Ichthyornithes subgroup --> Gulls, Skimmers (Charadriiformes/Lari) AND
- An Ichthyornithes subgroup --> Petrels (Procellariiformes), Sphenisciformes, Pelecaniformes
http://upload.wikimedia.org/wikipedia/commons/thumb/8/89/Iberomesornis-model.jpg/250px-Iberomesornis-model.jpg
Ichthyornis
http://upload.wikimedia.org/wikipedia/commons/thumb/f/f2/Ichthyornis_BW.jpg/250px-Ichthyornis_BW.jpg
Gull
http://upload.wikimedia.org/wikipedia/commons/thumb/1/13/Black-headed_Gull_-_St_James%27s_Park%2C_London_-_Nov_2006.jpg/250px-Black-headed_Gull_-_St_James%27s_Park%2C_London_-_Nov_2006.jpg
WADERS/SHOREBIRDS
- A Dromaeosaurid* subgroup -->
- An Enantiornithes shorebird subgroup --> Primitive shorebird (eg. Graculavus) --> Modern shorebirds - eg. plovers, oystercatchers, sandpipers (Charadriiformes/Charadrii)
http://upload.wikimedia.org/wikipedia/commons/thumb/8/89/Iberomesornis-model.jpg/250px-Iberomesornis-model.jpg
Shorebird
http://upload.wikimedia.org/wikipedia/commons/thumb/0/00/Calidris-pusilla-001.jpg/240px-Calidris-pusilla-001.jpg
AQUATIC BIRDS (Hesperornithes line)
- A Dromaeosaurid* subgroup -->
- An Enantiornithes aquatic subgroup --> Baptornithidae (Hesperornithes) --> (primarily foot-propelled) WEB FOOT diving bird orders, eg. Cormorants (Phalacrocoracidae), Loons (Gaviidae)). AND
- An Enantiornithes aquatic subgroup --> Hesperornithidae (Hesperornithes) --> (primarily foot-propelled) LOBE FOOT diving bird orders eg. Grebes (Podicipedidae).
Enantiornithes
http://upload.wikimedia.org/wikipedia/commons/thumb/8/89/Iberomesornis-model.jpg/250px-Iberomesornis-model.jpg
Hesperonis
http://upload.wikimedia.org/wikipedia/commons/thumb/d/d1/Hesperornis_BW.jpg/220px-Hesperornis_BW.jpg
Loon
http://bwfov.typepad.com/birders_world_field_of_vi/images/2007/07/18/loon_2.jpg
WATERFOWL (Anatidae)
- A Dromaeosaurid* subgroup -->
- An Enantiornithes waterbird subgroup --> Presbyornithids--> Modern Anseriformes (eg. Duck, Geese , Swan)
http://upload.wikimedia.org/wikipedia/commons/thumb/8/89/Iberomesornis-model.jpg/250px-Iberomesornis-model.jpg
Presbyornis
http://critters.pixel-shack.com/WebImages/crittersgallery/Presbyornis.jpg
Goose
http://upload.wikimedia.org/wikipedia/commons/thumb/c/c0/Magpie.goose.grooming.arp.750pix.jpg/222px-Magpie.goose.grooming.arp.750pix.jpg
LANDBIRDS (General)
- A Dromaeosaurid* subgroup --> Primitive flying birds (Enantiornithes) --> Modern Land Bird Orders
http://upload.wikimedia.org/wikipedia/commons/thumb/8/89/Iberomesornis-model.jpg/250px-Iberomesornis-model.jpg
Modern Land Bird
http://upload.wikimedia.org/wikipedia/commons/thumb/3/32/House_sparrow04.jpg/220px-House_sparrow04.jpg
LANDFOWL
- A Dromaeosaurid* subgroup --> Primitive flying birds (Enantiornithes) --> Modern "Galliformes" (eg. Chicken, Turkey, Pheasant, Quail) and Tinamiformes (Tinamou)
Enantiornithes
http://upload.wikimedia.org/wikipedia/commons/thumb/8/89/Iberomesornis-model.jpg/250px-Iberomesornis-model.jpg
Fowl
http://upload.wikimedia.org/wikipedia/commons/thumb/7/79/Sonnerathuhn.jpg/240px-Sonnerathuhn.jpg
Tinamou
http://leesbirdblog.files.wordpress.com/2010/06/red-winged-tinamou-rhynchotus-rufescens-by-c2a9wiki.jpg
LANDBIRDS (OWL)
- A Dromaeosaurid* subgroup -->
- Troodontid subgroup -->
- Primitive owls (eg. The supposed "Cretaceous owls"Bradycneme and Heptasteornis) -->
- Strigiformes (eg. owls) AND
- Caprimulgiformes (eg. nighthawks)
http://upload.wikimedia.org/wikipedia/commons/thumb/8/8f/Meilong_mmartyniuk_wiki.png/250px-Meilong_mmartyniuk_wiki.png
Owl
http://upload.wikimedia.org/wikipedia/commons/thumb/1/14/Northern_Spotted_Owl.USFWS-thumb.jpg/220px-Northern_Spotted_Owl.USFWS-thumb.jpg
*Dromaeosaurids developed from pterosaurs.
ANALYSIS
So far in the analysis, we have separate primitive bird lines leading to specific modern bird orders.
All the lines trace back to Dromaeosauridae (which then traces back to Pterosauria).
The primitive bird lines are based on:
- Ornithomimosaurids
- Enantiornithes
- Ichthyornithes
- Hesperornithes
- Troodontids
- Presbyornithids




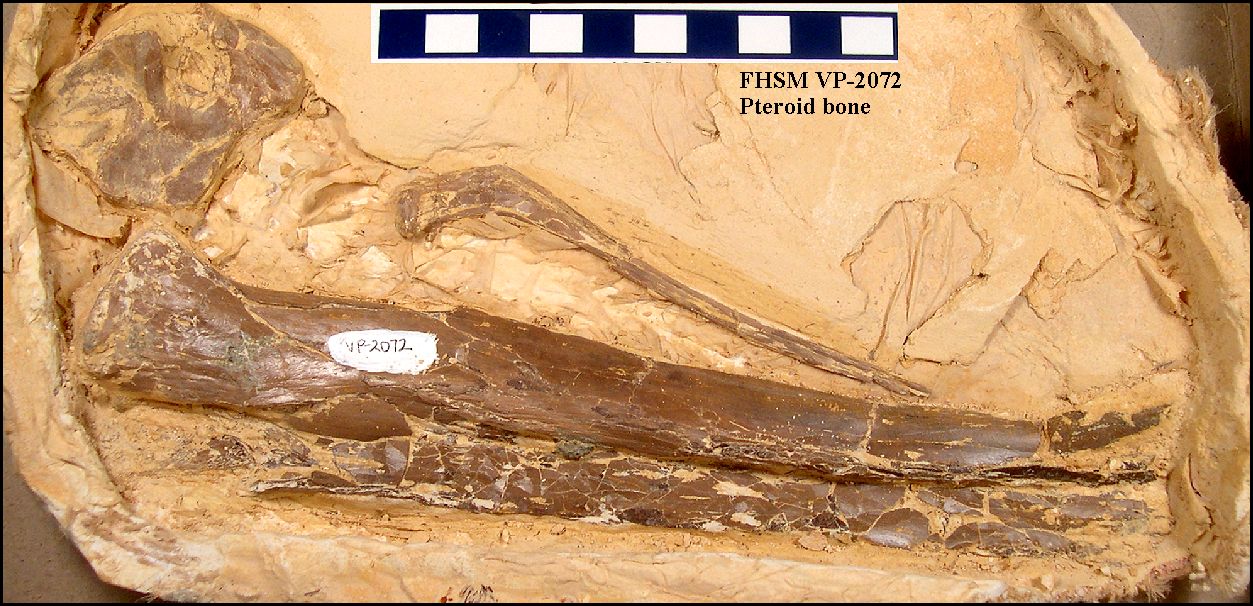




27%5B881:aafotp%5D2.0.co;2/production/images/large/i0272-4634-27-4-881-f05.jpeg)
27%5B881:aafotp%5D2.0.co;2/production/images/large/i0272-4634-27-4-881-f03.jpeg)

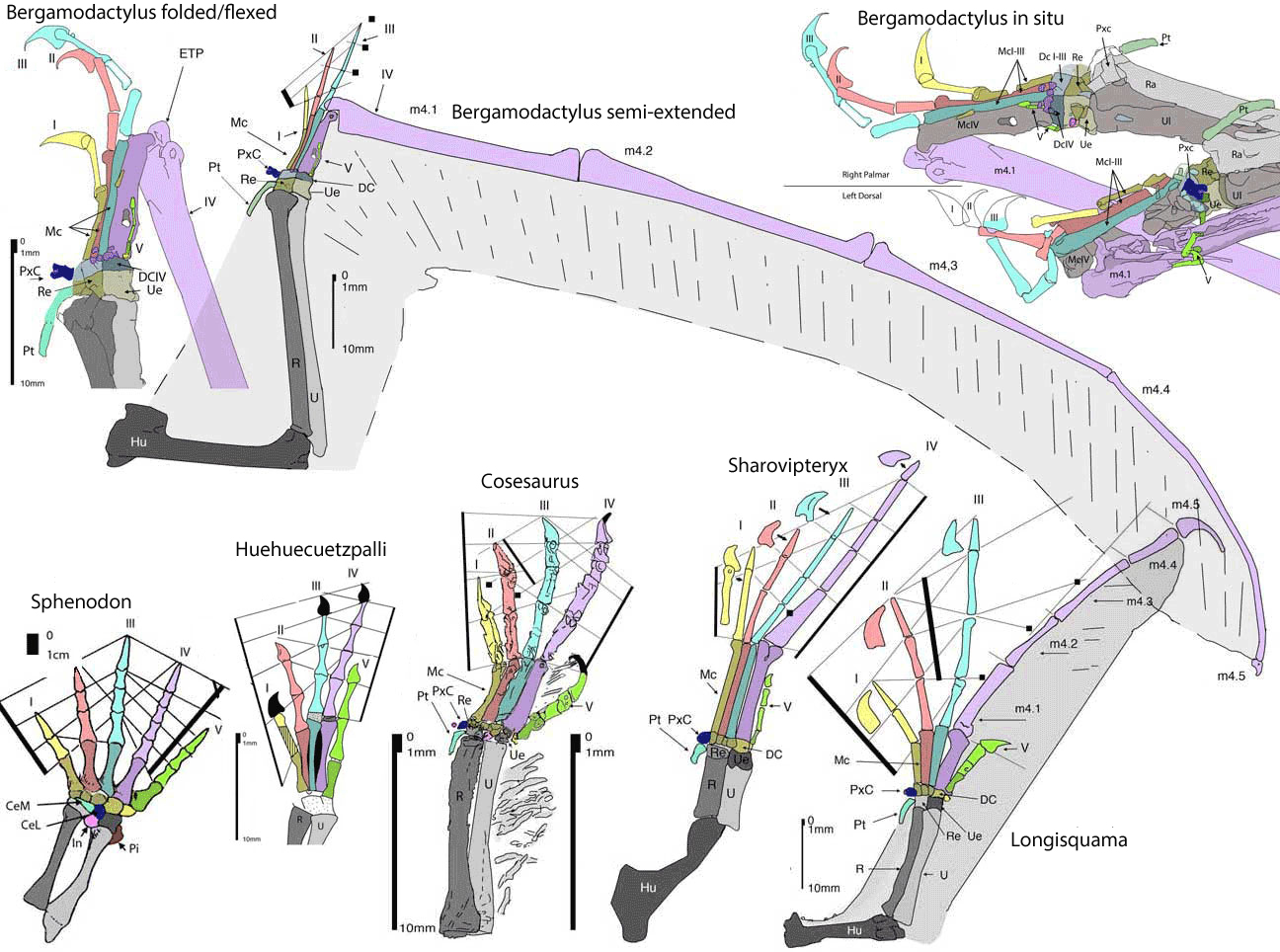


27%5B881:aafotp%5D2.0.co;2/production/images/medium/i0272-4634-27-4-881-f02.gif)

27%5B881:aafotp%5D2.0.co;2/production/images/medium/i0272-4634-27-4-881-f01.gif)















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}