The mainstream dinosaur-to-bird thinking is that Paraves is a node on a proposed lineage from dinosaur to bird. And that basal Paraves was ground-based and that the ground based Oviraptorosauria was a sister taxon.
However, the evidence is substantive that basal Paraves* was a power-flying, primitive bird. It had numerous flying bird-like characteristics. In addition, the evidence is substantive that Oviraptors were secondarily flightless descendants of basal Paraves, having lost/reduced their flying bird-like characteristics as they adapted secondarily to life on the ground, like an ancient Ostrich.
1. Mainstream dinosaur to bird hypothesis
The mainstream dinosaur to bird hypothesis is that Paraves was ground-based. And that Oviraptors were sister taxa to Paraves.

Paraves is a branch-based clade defined to include all dinosaurs which are more closely related to birds than to oviraptorosaurs. The ancestral paravian is a hypothetical animal: the first common ancestor of birds, dromaeosaurids, and troodontids which was not also ancestral to oviraptorosaurs. Little can be said with certainty about this animal. But the work of Xu et al. (2003), (2005) and Hu et al. (2009) provide examples of basal and early paravians with four wings,[11][12][13] adapted to an arboreal lifestyle who would only lose their hindwings when some adapted to a life on the ground and when avialans evolved powered flight.[14] Newer research also indicates that gliding, flapping and parachuting was another ancestral trait of Paraves, while true powered flight only evolved once, in the lineage leading to modern birds.[15]
Oviraptorosaurs ("egg thief lizards") are a group of feathered maniraptoran dinosaurs from the Cretaceous Period of what are now Asia and North America. They are distinct for their characteristically short, beaked, parrot-like skulls, with or without bony crests atop the head. They ranged in size from Caudipteryx, which was the size of a turkey, to the 8 metre long, 1.4 ton Gigantoraptor.[4] The group (along with all maniraptoran dinosaurs) is close to the ancestry of birds. Analyses like those of Maryanska et al (2002) and Osmólska et al. (2004) suggest that they may represent primitive flightless birds.[5][6]
Paraves bird-like characteristics

Paraves bird-like characteristics (including the Pennaraptora bird-like characteristics)
V-shaped furcula; ACVP, advanced costosternal ventilator pump; AFC, initial arm flapping capability; BMR, further increased basal metabolic rate; CE, cerebral expansion; ES, increased egg size; IL, iterative laying; LFC, increased laterally folding capability; MO, monoautochronic ovulation; PC, paternal care; PPO, partial pubis posterior orientation; SAE, slightly asymmetrical egg; SBT*, short bony tail; TFH, three-fingered hand; VF, symmetrical vaned feathers.* NOTE: Only highly derived members (Pygostylia) had a short bony tail. Basal Pennaraptora had a long bony tail.
AET, arm elongation and thickening; AL, initial aerial locomotion; EM, extreme miniaturization; IA, egg with increased asymmetry; ICI, improved contact incubation; KBL, partial knee-based locomotion; LP, eggshell with low porosity; TL, eggshell with third (external) layer; US, egg with unornamented surface; VABRE, visually associated brain regions elaboration; VF*, asymmetrical vaned feathers.* Note: Only derived members of Paraves had asymmetrical vaned feathers. Basal Paraves had symmetrical vaned feathers.
Several flight-related anatomical features, such as hollow bones and the furcula, originated in early theropods; basal paravians had many hallmark features necessary for flight, including extremely small body size (50, 70); a laterally oriented, long, and robust forelimb (22, 67); an enlarged forebrain and other derived neurological adaptations (71); and large flight feathers (Figs. 1 and 2). Particularly surprising are the recent discoveries of large flight feathers forming a planar surface on the legs of some basal paravians—for example, those with asymmetrical vanes on both the tibia and metatarsus of some basal dromaeosaurs, such as Microraptor (59); large feathers with symmetrical vanes on both the tibia and metatarsus of the troodontid Anchiornis (45), the basal bird Sapeornis, and several other basal paravians (135); and large vaned feathers on tibiae of several basal birds including Archaeopteryx, confuciusornithids, and enantiornithines (135). These structures clearly would have been relevant to flight origins.
For example, the unusually crouched hindlimb for bipedal locomotion that characterizes modern birds was acquired in stepwise fashion through much of theropod evolution (67), and both the furcula (68) and the “semilunate” carpal (69) appeared early in theropod evolution. Notably, major bird characteristics often exhibit a complex, mosaic evolutionary distribution throughout the theropod tree, and several evolutionary stages are characterized by accelerated changes (70). For example, the early evolution of paravian theropods features cerebral expansion and elaboration of visually associated brain regions (71), forelimb enlargement (22, 67), acquisition of a crouched, knee-based hindlimb locomotor system (67), and complex pinnate feathers associated with increased melanosome diversity, which implies a key physiological shift (72). Together these features may suggest the appearance of flight capability at the base of the Paraves (22, 67).Characteristics:
1.1 Tetrapteryx (4 winged)
It has been hypothesized that bird flight went through a four-winged ("tetrapteryx") stage.
Pascal Godefroit et al (2013a)
Epidendrosaurus, Epidexipteryx and Eosinopteryx, also from the Middle–Late Jurassic of northeastern China, are here regarded as basal, non-eumaniraptoran paravians. Thus our phylogeny is entirely consistent with the presence of a tetrapterygian condition (= four winged) and elongated rectrices in basal eumaniraptorans.
Dongyu Hu et al (2009)
The early evolution of the major groups of derived non-avialan theropods is still not well understood, mainly because of their poor fossil record in the Jurassic. Here we report on an exceptionally well-preserved small theropod specimen [Anchiornis] collected from the earliest Late Jurassic Tiaojishan Formation of western Liaoning, China2. The specimen is referable to the Troodontidae, which are among the theropods most closely related to birds. .... the extensive feathering of this specimen, particularly the attachment of long pennaceous feathers to the pes, sheds new light on the early evolution of feathers and demonstrates the complex distribution of skeletal and integumentary features close to the dinosaur–bird transition.
Large pennaceous feathers are now known to occur on the lower leg and particularly the metatarsus of at least one basal member of each of the three major paravian groups, namely the basal troodontid Anchiornis, the basal avialan Pedopenna and the basal dromaeosaurid Microraptor13,16. Furthermore, many basal avians have proportionally large pennaceous feathers on the lower leg13,17, which are reduced in more derived birds. This suggests that large pennaceous feathers first evolved distally on the hindlimbs, as on the forelimbs and tail. This distal-first development led to a four-winged condition at the base of the Paraves. Whereas the large feathers of the forewing developed further in subsequent avian evolution, the large hindwing feathers were reduced and even lost12. This suggests that extensive feathering of the pes was a critical modification in the transition to birds and that the pedal scales of extant birds might be secondarily derived structures, a possibility also supported by some developmental studies18
Tetrapterygidae (meaning "four-wings") is a group of four-winged dinosaurs proposed by Sankar Chatterjee in the second edition of his book The Rise of Birds: 225 Million Years of Evolution, where he included Microraptor, Xiaotingia, Aurornis, and Anchiornis.[1] The group was named after the characteristically long flight feathers on the legs of all included species, as well as the theory that the evolution of bird flight may have gone through a four-winged (or "tetrapteryx") stage, first proposed by naturalist William Beebe in 1915.[2] Chatterjee suggested that all dinosaurs with four wings formed a natural group exclusive of other paravians, and that this family was the sister taxon to the group Avialae, although most phylogenetic analyses have placed the animals of his Tetrapterygidae elsewhere in Paraves, such as Xiaotingia, Aurornis, and Anchiornis being placed in Avialae.[3]
1.2 Arboreal
https://en.wikipedia.org/wiki/Scansoriopterygidae
Scansoriopteryx heilmanni (and its likely synonym Epidendrosaurus ninchengensis) was the first non-avian dinosaur found that had clear adaptations to an arboreal or semi-arboreal lifestyle–it is likely that they spent much of their time in trees.Xing Xu et al (2005)
The unusual presence of long pennaceous feathers on the feet of basal dromaeosaurid dinosaurs has recently been presented as strong evidence in support of the arboreal–gliding hypothesis for the origin of bird flight, but it could be a unique feature of dromaeosaurids and thus irrelevant to the theropod–bird transition. Here, we report a new eumaniraptoran theropod [Pedopenna] from China, with avian affinities, which also has long pennaceous feathers on its feet. This suggests that such morphology might represent a primitive adaptation close to the theropod–bird transition. The long metatarsus feathers are likely primitive for Eumaniraptora and might have played an important role in the origin of avian flight.
1.3 Feathers
https://en.wikipedia.org/wiki/Anchiornis
While the first specimen of Anchiornis preserved only faint traces of feathers around the preserved portion of the body, the well-preserved second specimen showed nearly complete feather preservation, allowing researchers to identify the structure of the feathers and how they were distributed.
1.4 Long and robust forelimbs
Xing Xu et al (2011)
1.5 Miniaturization
Archaeopteryx is widely accepted as being the most basal bird, and accordingly it is regarded as central to understanding avialan origins; however, recent discoveries of derived maniraptorans have weakened the avialan status of Archaeopteryx. Here we report a new Archaeopteryx-like theropod from China. This find further demonstrates that many features formerly regarded as being diagnostic of Avialae, including long and robust forelimbs, actually characterize the more inclusive group Paraves (composed of the avialans and the deinonychosaurs).
1.5 Miniaturization
Michael J. Benton (2015)
These studies of bird origins [56–59] used different datasets, different phylogenies, and different analytical techniques, and yet they converged on the same result. As an example, Puttick et al. [56] showed that miniaturization and wing expansion, critical anatomical requirements to be a bird, arose some 10 Myr before Archaeopteryx among the wider clade Paraves (figure 4), and that the rate of change was 160 times the normal evolutionary rate, suggesting a rapid, adaptive switch that enabled the diversification and success of this clade of tiny, possibly tree-climbing and gliding dinosaurs.
1.6 Pelvic limb
Vivian Allen et al (2013)
Rather than a discrete transition from more-upright postures in the basal-most birds (Avialae) and their immediate outgroup deinonychosauria5,6, our results support hypotheses of a gradual, stepwise acquisition of more-crouched limb postures across much of theropod evolution1–4, although we find evidence of an accelerated change within the clade Maniraptora (birds and their closest relatives, such as deinonychosaurs). In addition, whereas reduction of the tail is widely accepted to be the primary morphological factor correlated with centre-of-mass position and, hence, evolution of hindlimb posture1, 2, 3, 4, 5, 6, 7, 8, we instead find that enlargement of the pectoral limb and several associated trends have a much stronger influence. Intriguingly, our support for the onset of accelerated morpho-functional trends within Maniraptora is closely correlated with the evolution of flight. Because we find that the evolution of enlarged forelimbs is strongly linked, via whole-body centre of mass, to hindlimb function during terrestrial locomotion, we suggest that the evolution of avian flight is linked to anatomical novelties in the pelvic limb as well as the pectoral.
1.7 Brain
Amy M. Balanoff et al (2013)
The brain of Archaeopteryx is not volumetrically avian but conforms to the expectation of a generalized paravian. Thus, if Archaeopteryx has a ‘flight-ready’ brain3 , which is almost certainly the case given its postcranial morphology, then so did other paravians. The hypothesis that dromaeosaurs and troodontids had the neurological capabilities required of powered flight, gliding, or some intermediate condition is congruent with the discovery of the ‘four-winged’ deinonychosaurs, Microraptor zhaoianus20 and Anchiornis huxleyi5,6
There are a number of problems with the mainstream idea that Paraves was ground dwelling. Problems include:
- flight had to evolve multiple times independently (homoplasy).
- it requires numerous exaptations (they were not using their bird-like characteristics for flying)
- the timing of found fossils is inconsistent with it (stratigraphic incongruence)
- character optimization does not favor that alternative
- it requires an implausible rate of evolution
- the hindwing feathers would interfere with running
2.1 Extensive Homoplasies
Many biologists aim to avoid homoplasies in grouping species together and therefore it is frequently a goal to eliminate groups that are found to be polyphyletic. This is often the stimulus for major revisions of the classification schemes.
Convergent evolution is the independent evolution of similar features in species of different lineages. Convergent evolution creates analogous structures that have similar form or function, but that were not present in the last common ancestor of those groups.[1] The cladistic term for the same phenomenon is homoplasy
A polyphyletic group is characterized by one or more homoplasies: phenotypes which have converged or reverted so as to appear to be the same but which have not been inherited from common ancestors.
The early history of flight is highly complex. As Longrich and colleagues [8] point out, flight capability is likely to have evolved independently on multiple occasions among Archaeopteryx and its kin. For example, flight feathers with asymmetrical vanes seem to have evolved independently at least twice near the dinosaur-bird transition, once near the base of the Avialae and once in the deinonychosaurs [8].
As a result of the high amount of homoplasy that characterizes derived maniraptoran evolution, the identity of the avian sister taxon remains debated despite the rapid accumulation of morphological data.
Xiaotingia zhengi independently evolved some salient features seen in other maniraptoran taxa, which highlights the extensive homoplasy that exists among maniraptorans.
Pascal Godefroit et al (2013b)
Note, however, that this phylogeny remains only weakly supported presumably due to the numerous homoplasies widely distributed across coelurosaurian phylogeny3,17
Amy M. Balanoff et al (2013)
Features that were once considered exclusive to modern birds, such as feathers and a furcula, are now known to have first appeared in non-avian dinosaurs1 . However, relatively little is known of the early evolutionary history of the hyperinflated brain that distinguishes birds from other living reptiles and provides the important neurological capablities required by flight2 . Here we use high-resolution computed tomography to estimate and compare cranial volumes of extant birds, the early avialan Archaeopteryx lithographica, and a number of non-avian maniraptoran dinosaurs that are phylogenetically close to the origins of both Avialae and avian flight. Previous work established that avian cerebral expansion began early in theropod history and that the cranial cavity of Archaeopteryx was volumetrically intermediate between these early forms and modern birds3,4. Our new data indicate that the relative size of the cranial cavity of Archaeopteryx is reflective of a more generalized maniraptoran volumetric signature and in several instances is actually smaller than that of other non-avian dinosaurs. Thus, bird-like encephalization indices evolved multiple times, supporting the conclusion that if Archaeopteryx had the neurological capabilities required of flight, so did at least some other non-avian maniraptorans. This is congruent with recent findings that avialans were not unique among maniraptorans in their ability to fly in some form5,6.
This indicates that the origin of flight in avialans was more complex than previously thought and might have involved several convergent achievements of aerial abilities.
2.2. Numerous Exaptations
In the dinosaur to bird theory, there is an extensive number of required exaptations.
Exaptation and the related term co-option describe a shift in the function of a trait during evolution. For example, a trait can evolve because it served one particular function, but subsequently it may come to serve another.
Abducted wrists, feathers, enlarged brains and laterally oriented, long and robust forelimbs are claimed to have evolved before they were used for flight.
ABDUCTED WRISTS (Semilunate carpal)
ABDUCTED WRISTS (Semilunate carpal)
Corwin Sullivan et al (2010)
Carpal asymmetry [abducted wrists] would have permitted avian-like folding of the forelimb in order to protect the plumage, an early advantage of the flexible, asymmetric wrist inherited by birds.
However, it is likely that mobility of the wrist was initially associated with other functions, such as predation (Padian 2001).
Partial folding of the wing during the upstroke in extant birds, which requires significant abduction of the wrist (Vazquez 1992) could then be seen as an exaptation of a capability that originally evolved in non-volant, non-avialan maniraptorans.
It had originally been proposed that this [wrist] flexibility could be attributed to hunting, but the same changes are seen in maniraptorans that were herbivores and omnivores so it is unlikely that hunting provides the answer. Instead, the authors of the new study propose, the ability to fold the hands backwards would have protected the feathers of the arms. This would have prevented the feathers from getting damaged or from being in the way as the dinosaurs moved about, although the authors recognize that this hypothesis requires further evidence.
Perhaps more significant, however, is how this wing-folding mechanism may have allowed birds to take to the air. Birds do flex their wrists while flapping their wings to fly, and so it appears that the wrist flexibility that first evolved in dinosaurs was later co-opted for flight in birds. This is what is known as "exaptation," or when a previous adaptation takes on a new function. Indeed, as more is discovered about the evolution of birds, the more traits paleontologists find that evolved for one function but have been co-opted for another at a later point (feathers themselves being the most prominent example). There is relatively little separating birds from their feathered dinosaur ancestors.
As Darwin elaborated in the last edition of The Origin of Species,[14] many complex traits evolved from earlier traits that had served different functions. By trapping air, primitive wings would have enabled birds to efficiently regulate their temperature, in part, by lifting up their feathers when too warm. Individual animals with more of this functionality would more successfully survive and reproduce, resulting in the proliferation and intensification of the trait.
Eventually, feathers became sufficiently large to enable some individuals to glide. These individuals would in turn more successfully survive and reproduce, resulting in the spread of this trait because it served a second and still more beneficial function: that of locomotion. Hence, the evolution of bird wings can be explained by a shifting in function from the regulation of temperature to flight.
Exaptation is a term used in evolutionary biology to describe a trait that has been co-opted for a use other than the one for which natural selection has built it.
It is a relatively new term, proposed by Stephen Jay Gould and Elisabeth Vrba in 1982 to make the point that a trait’s current use does not necessarily explain its historical origin. They proposed exaptation as a counterpart to the concept of adaptation.
For example, the earliest feathers belonged to dinosaurs not capable of flight. So, they must have first evolved for something else. Researchers have speculated early feathers may have been used for attracting mates or keeping warm. But later on, feathers became essential for modern birds’ flight.
Gareth Dyke et al (2013)
This further supports the hypothesis that "flight feathers" that first evolved in dinosaurs for non-aerodynamic functions were later adapted to form lifting surfaces.[15]
Several ancient dinosaurs evolved the brainpower needed for flight long before they could take to the skies, scientists say.
Bird brains tend to be more enlarged compared to their body size than reptiles, vital for providing the vision and coordination needed for flight.Scientists using high-resolution CT scans have now found that these "hyper-inflated" brains were present in many ancient dinosaurs, and had the neurological hardwiring needed to take to the skies. This included several bird-like oviraptorosaurs and the troodontids Zanabazar junior, which had larger brains relative to body size than that of Archaeopteryx.
The brain of Archaeopteryx is not volumetrically avian but conforms to the expectation of a generalized paravian. Thus, if Archaeopteryx has a ‘flight-ready’ brain3 , which is almost certainly the case given its postcranial morphology, then so did other paravians. The hypothesis that dromaeosaurs and troodontids had the neurological capabilities required of powered flight, gliding, or some intermediate condition is congruent with the discovery of the ‘four-winged’ deinonychosaurs, Microraptor zhaoianus20 and Anchiornis huxleyi5,6
LATERALLY ORIENTED, LONG AND ROBUST FORELIMBS
Forelimb myology and breathing apparatusSeveral flight-related anatomical features, such as hollow bones and the furcula, originated in early theropods; basal paravians had many hallmark features necessary for flight, including extremely small body size (50, 70); a laterally oriented, long, and robust forelimb (22, 67); an enlarged forebrain and other derived neurological adaptations (71); and large flight feathers (Figs. 1 and 2). Particularly surprising are the recent discoveries of large flight feathers forming a planar surface on the legs of some basal paravians—for example, those with asymmetrical vanes on both the tibia and metatarsus of some basal dromaeosaurs, such as Microraptor (59); large feathers with symmetrical vanes on both the tibia and metatarsus of the troodontid Anchiornis (45), the basal bird Sapeornis, and several other basal paravians (135); and large vaned feathers on tibiae of several basal birds including Archaeopteryx, confuciusornithids, and enantiornithines (135). These structures clearly would have been relevant to flight origins.
Christian Foth et al (2014)
The current results show that pennaceous feathers were present in the common ancestor of Pennaraptora. As in extant birds, this feather type probably fulfilled several biological roles related to body insulation, manoeuvrability, brooding, camouflage and display7,16. However, giventhe great diversity of pennaceous feathers found within different body regions and across the phylogeny, it seems plausible that the evolution of this feather type (especially in the wing, hindlimbs and tail) was primarily driven by display functions. These structures then represented an exaptation that was repeatedly, and probably convergently, recruited for aerodynamic functions. This indicates that the origin and evolution of flight in theropod dinosaurs were more complex than previously thought, and (as already shown for other anatomical complexes, for example forelimb myology and breathing apparatus) could draw on structures that evolved in different functional contexts.
OTHER
Placed in context of avian evolution, the grasping foot of Deinonychus and other terrestrial predatory paravians is hypothesized to have been an exaptation for the grasping foot of arboreal perching birds. Here we also describe “stability flapping”, a novel behaviour executed for positioning and stability during the initial stages of prey immobilisation, which may have been pivotal to the evolution of the flapping stroke. These findings overhaul our perception of predatory dinosaurs and highlight the role of exaptation in the evolution of novel structures and behaviours.
Did preadaptations for flight precede the origin of birds (Aves)? The origin of flight in birds is one of the great evolutionary transitions and has received considerable attention in recent years (Padian and Chiappe 1998; Clarke and Middleton 2008; Dececchi and Larrson 2009; Benson and Choiniere 2013; Dececchi and Larrson 2013). The evolution of birds is often considered coincident with the origins of flight, but many traits associated with flight evolved before the origin of Aves (Padian and Chiappe1998).
Gregory M. Erickson et al (2009)
This suggests that the initial conquest of the air was achieved using lower metabolic rates than are characteristic of today's avian flyers. It appears that the closest non-avialan relatives of birds were physiologically preadapted for powered flight and only anatomical adaptations were involved when birds first ventured into the air.
Frances C. James et al (2009)
Although some or all of the flight-related characters of birds, including those found in the maniraptorans, may have evolved for purposes other than flight, current exaptational explanations offered by BMT proponents are often not fully formulated and rarely offer a biologically plausible hypothesis to account for their origin (Feduccia 1985, 1993, 1995; Paul 2002; Feduccia et al. 2005, 2007). Under such conditions, exaptational explanations should not be regarded as having priority (Rose and Lauder 1996), and adaptational accounts should not be discarded. If the most birdlike maniraptoran clades belong within Aves, problematic exaptational explanations, including those for the origin of flight feathers, are unnecessary.
2.3. Stratigraphic Incongruence
Earlier analyses had placed Oviraptors earlier than Paraves. However, the Oviraptor fossils that have been found, are actually tens of millions of years later than Paraves. So a claim of lengthy ghost lineages is required.
Scansoriopterygids (Temporal range: Late Jurassic, 165–156 Ma)
Anchiornis (Temporal range: Late Jurassic, 161–160.5 Ma)
Aurornis (Temporal range: Late Jurassic, 160 Ma)
Xiaotingia (Temporal range: Late Jurassic, 160 Ma)
Pedopenna (Temporal range: Middle or Late Jurassic, 164 Ma)
Oviraptors (Temporal range: Cretaceous, 130-66 Ma)
2.4. Character Optimization
Character optimization is the process by which alternative reconstructions of a character on a cladogram are evaluated.
Ingi Agnarsson et al (2008)
Ingi Agnarsson et al (2008)
Character optimization is the process by which alternative reconstructions of a character on a cladogram are evaluated.
De Pinna (1991, p. 386) argued that ‘‘… absences stand at a lower ontological level as observations, when compared to presences (Nelson and Platnick, 1981: 29; Patterson, 1982: 30).’’ In other words, there can be asymmetry in the information content of primary homology statements of character states. Complex features share detailed similarities strengthening the conjecture of homology between them, whereas absences are a weaker form of primary homology statements (see also Rieppel and Kearney, 2002). Therefore, a theoretical basis may exist for favouring some equally parsimonious optimizations over others. In the absence of compelling evidence to the contrary, ambiguous optimization is better resolved in favour of secondary losses (reversals) over parallel gains of complex structures. This is more consistent with the stronger conjecture of homology based on observable detailed similarity, rather than mere absence (De Pinna, 1991); complexity tests similarity (Agnarsson and Coddington, 2008; see also Richter, 2005; Scholtz, 2005; Agnarsson et al., 2007). When characters lack asymmetry in character state complexity (e.g. the states ‘‘red’’ and ‘‘blue’’), little, if any, grounds exist to favour one optimization over another (Richter, 2005).
Zhonghe Zhou (2014)
However, contrary to Foth and colleagues [11], it is probably premature to reject a four-winged stage during the origin of avian flight. The leg feathers of Archaeopteryx are relatively short, yet they are weakly curved like the flight feathers and are not so short that an aerodynamic function can be excluded. There is still no evidence showing unequivocally that leg feathers lacked aerodynamic function in the direct ancestor of birds. In fact, Microraptor probably represents only one of many extinct feathered dinosaurs that maintained an ancestral aerodynamic role in the leg feathers [2]. A recent study on the leg feathers of a primitive bird, Sapeornis, suggested a distal-to-proximal pattern of reduction during leg feather evolution [13]. It is possible that an aerodynamic function of pennaceous feathers could have evolved several times in various theropod lineages; however, it is also more likely that flight and aerodynamic functions of the pennaceous feathers could have been lost many times in theropod (including birds) evolution — during evolution, the loss of features is far more common than the evolution of novel features.
Mesozoic Birds: Above the Heads of Dinosaurs
See page 442.
Taxa on terminal branches have novelties (autapomorphies) not present in the lineal ancestors of birds. Including autapomorphies in a reconstructed ancestor is therefore unjustified and a sign that its characters were not optimized.
Valentin Rineau1 , Anaïs Grand2 , René Zaragüeta3 , Michel Laurin1, 4 (2015)
Experimental systematics: sensitivity of cladistic methods to polarization and character ordering schemes
We discuss implications of these points of view compared to the assumption inherent in 3ta (i.e., that reversals should not support a clade as other synapomorphies do) on evolutionary models.
A particularly controversial issue in cladistics concerns the treatment of reversals.
We take this opportunity to propose a nomenclatural clarification about reversals (based on the example in Fig. 3A) as secondary homology hypotheses; thus, this clarification is valid both for parsimony and for 3ta. First rounds of reversals are generally called ‘secondary losses’ (e.g. (Carine and Scotland, 1999), when in fact, only the absence should be considered secondary and the loss in itself should be considered as an event that appeared for the first time (i.e., primary). Thus, a character state is primitively absent (primary absence; state 0 on Fig. 3). It can then appear; this is a primary appearance (of state 1), denoted +1 on Fig. 3A. It can be subsequently lost (-1, reversal to state 0, but identified as 0* on Fig. 3a, for greater clarity); this should be called a primary loss, which results in a secondary absence. After this, a secondary gain (+2) can lead to secondary presence (1* in Fig. 3A), and a secondary loss (-2) can lead to ternary absence (0** in Fig. 3), etc.
2.5. Implausible Rates of Evolution
Mark N Puttick et al (2014)
Before the origin of Aves, on the branch leading to Paraves, high rates of evolution led to a smaller body size and a relatively larger forelimb in Paraves. These changes are on a single branch leading to Paraves, representing a shift to a new smaller size and larger forelimb at this point. Rapid miniaturization and relative forelimb elongation at the root of Paraves explain the similarities between early birds and other Paraves (Padian and Chiappe 1998; Turner et al. 2007; Clarke and Middleton 2008; Novas et al. 2012). As with previous studies, we find strong evidence for paravian miniaturization (Xu et al. 2003; Turner et al. 2007; Novas et al. 2012) and no trend for forelimb elongation (Dececchi and Larrson 2013), but the identification here of increased rates of evolution of size-dependent forelimb and body size at the origin of Paraves emerges from our novel analytical approach. As with a recent study (Deccechi and Larrson 2013), we find evidence for a different allometric relationship between forelimb size and body size in Aves, but this result is altered by different phylogenetic topologies, and we find little evidence for elevated rates leading to or within Aves.
Michael S. Y. Lee et al (2014)
Recent discoveries have highlighted the dramatic evolutionary transformation of massive, ground-dwelling theropod dinosaurs into light, volant birds. Here, we apply Bayesian approaches (originally developed for inferring geographic spread and rates of molecular evolution in viruses) in a different context: to infer size changes and rates of anatomical innovation (across up to 1549 skeletal characters) in fossils. These approaches identify two drivers underlying the dinosaur-bird transition. The theropod lineage directly ancestral to birds undergoes sustained miniaturization across 50 million years and at least 12 consecutive branches (internodes) and evolves skeletal adaptations four times faster than other dinosaurs. The distinct, prolonged phase of miniaturization along the bird stem would have facilitated the evolution of many novelties associated with small body size, such as reorientation of body mass, increased aerial ability, and paedomorphic skulls with reduced snouts but enlarged eyes and brains.

The ancestors of Paraves first started to shrink in size in the early Jurassic 200 million years ago, and fossil evidence show that this theropod line evolved new adaptations four times faster than other groups of dinosaurs,[8] and was shrinking 160 times faster than other dinosaur lineages were growing.[9]
Daniel T. Ksepka (2014)
Rates of morphological evolution can be quantified using time-calibrated phylogenetic trees. Brusatte et al. [1] employ the new phylogeny as the framework for conducting likelihood analyses of skeletal character evolution. These tests recover high rates of evolution both within the bird clade and also along a series of nodes on the theropod ‘backbone’ leading to birds. In agreement with these results, a recent Bayesian analysis using a different character dataset also found support for a sustained higher rate of skeletal evolution along this backbone [11], though the two studies disagree slightly on how deep in the tree the onset of higher rates occurs. A different set of tests in the Brusatte et al. [1] study compares rates between clades, revealing that birds as a clade exhibited a higher rate of skeletal evolution than other theropod clades. As a whole, these results suggest that birds are indeed a special case, leading to the hypothesis that the completion of the avian skeletal plan and development of powered flight opened the door to new ecological niches and triggered a burst of evolution [1].
Here again, the lineage leading to birds stands out as an exception, with maniraptoran theropods sustaining high rates of size evolution relative to other dinosaur lineages[12]. A third recent study [13] employing likelihood methods capable of detecting branch-specific rate shifts places the shift to higher rates of size evolution on the branch leading to Paraves (the clade uniting birds, dromaeosaurids, and troodontids).
2.6. The hindwing feathers would interfere with running
with large foot remiges cursorial locomotion was likely problematic for Anchiornis.
3.0 The Alternative
In order to analyze the alternative it is necessary to identify a new clade called "Euparaves". Euparaves is the node based version of Paraves. (For details see Appendix 4).
Euparaves (node-based) definition:
The last common ancestor of Epidendrosaurus ninchengensis (Scansoriopterygidae) and Passer domesticus (Aves) and all descendants thereof.
The alternative is that basal Euparaves was a flying primitive bird, that gave rise to other flying primitive birds and to secondarily-flightless ground-based creatures including the Oviraptors. "Secondarily flightless" means having descended from a flying ancestor.
The Oviraptors lost/reduced their flying bird-like characteristics when they settled on the ground and became secondarily flightless, like an ancient Ostrich. (For example the Oviraptors lost their leg feathers which would be a hindrance to running on the ground).
The hypothesis that Euparaves was a flying primitive bird is supported by the evidence and does not require the extensive amount of exaptation and convergence (homoplasy) that the current mainstream hypothesis requires. It is thus the more plausible and parsimonious hypothesis.
Oddly, this hypothesis has never been objectively analyzed.
Suggested phylogeny

4.0 Could Euparaves fly?
Since Euparaves contains the previously defined Paraves, we can attribute to Euparaves what we know about Paraves.
Basal Euparaves includes Anchiornis, Aurornis, Scansoripoterygids, Pedopenna, Xiaotingia and others.
We have seen the extensive set of bird characteristics they had. Are there reasons to think they could not fly?
We need to consider possible objections related to forewings, hindwings, airfoil, sternum, propatagium, semilunate carpal and feathers.
4.1. Forewings
It is sometimes thought that the primitive birds could not fly because they did not have the needed shoulder mechanism for powered flight. It is true that the lack of a derived supracoracoideus precluded takeoff from the ground. But there is no reason to think that powered-flight takeoff from tree branches was not possible.
Chatterjee and Templin also ruled out the possibility of a ground-based takeoff. Microraptor lacked the necessary adaptations in its shoulder joint to lift its front wings high enough vertically to generate lift from the ground, and the authors argued that a ground-based takeoff would have damaged flight feathers on the feet. This leaves only the possibility of launching from an elevated perch, and the authors noted that even modern birds do not need to use excess power when launching from trees, but use the downward-swooping technique they found in Microraptor.[4]
Feduccia (1999)
The dorsal elevators, principally the deltoideus major, can effect the recovery stroke by themselves, as they did in Archaeopteryx. The German anatomist Maxheinz J. Sy proved this when he cut the tendons of the supracoracoideus in living crows and pigeons (1936). Sy found that pigeons were capable of normal, sustained flight; the only capacity they lost was the ability to take off from level ground.
4.2. Hindwings
It is sometimes thought that the primitive birds lacked sufficient lift. But the hindwings generated additional lift.
In Microraptor, the metatarsal feathers are proportionally large with highly asymmetrical vanes (1), whereas in Pedopenna, Anchiornis, and Sapeornis they are proportionally smaller and have nearly symmetrical vanes (2, 3). In all cases, however, the metatarsal feathers are similar in general arrangement (nearly perpendicular to the metatarsus, forming a large flat surface) and in having stiff vanes and curved rachises. These features suggest that the metatarsal feathers were aerodynamic in function(12), providing lift, creating drag, and/or enhancing maneuverability, and thus played a role in flight (1–7, 24, 25). The presence of metatarsal feathers with a probable aerodynamic function in both deinonychosaurians and avialans strongly supports the interpretation that leg feathers were an important factor in the origin of avialan flight, although the nature of their biomechanical contribution to flight ability in taxa that possessed them is debated (1–7, 24–26).
The evolution of powered flight in birds remains a contentious issue in vertebrate paleontology. The diminutive predatory dinosaur Microraptor gui preserves evidence of extensive, lift-generating feathers on each manus and forearm, but also preserves evidence of lift-generating feathers associated with the hindlimbs, effectively forming a pair of “hindwings”.
4.3. Airfoil
It is sometimes thought that the primitive birds could not fly because they did not have asymmetric flight feathers and thus lacked an airfoil. It is true that they did not have asymmetric flight feathers. But that does not preclude flapping flight.
Nicholas R. Longrich et al (2012)
Here, we redescribe the wings of the archaic bird Archaeopteryx lithographica [3-5] and the dinosaur Anchiornis huxleyi [12, 13] and show that their wings differ from those of Neornithes in being composed of multiple layers of feathers. In Archaeopteryx, primaries are overlapped by long dorsal and ventral coverts. Anchiornis has a similar configuration but is more primitive in having short, slender, symmetrical remiges. Archaeopteryx and Anchiornis therefore appear to represent early experiments in the evolution of the wing. This primitive configuration has important functional implications: although the slender feather shafts of Archaeopteryx [14] and Anchiornis [12] make individual feathers weak, layering of the wing feathers may have produced a strong airfoil.
Teresa J. Feo et al (2015)
The elongated wing feathers of Mesozoic Paraves [Euparaves] exhibit small barb angles in cutting-edge leading vanes that are comparable with those of modern flying birds (figure 4b). This suggests that the leading vanes of these Mesozoic feathers are functionally similar to those of modern birds, and were similarly capable of withstanding aerodynamic forces in airflow. Furthermore, our observations document that the outer hindwing feathers of the four-winged dromaeosaurid Microraptor were similar in vane structure to the primary feathers of two-winged Mesozoic taxa, corroborating previous interpretations of their aerodynamic function [33]. The presence of small cutting-edge barb angles, in conjunction with sufficient vane asymmetry for feather pitch stability, support the conclusion that some form of aerial locomotion was plesiomorphic for the most exclusive clade including Microraptor and modern birds."4.4. Sternum
It is sometimes thought that the primitive birds could not fly because some did not have an ossified sternum. It is true that some did not have an ossified sternum but that does not preclude flapping flight.
STORRS L. OLSON et al (1979)
Furthermore, the supracoracoideus muscle, and hence an ossified sternum, is not necessary to effect the recovery stroke of the wing. Thus the main evidence for
Archaeopteryx having been a terrestrial, cursorial predator is invalidated. There is nothing in the structure of the pectoral girdle of Archaeopteryx that would preclude its having been a powered flier.
O'Connor et al (2014)
Ossified sternal plates (present and fused in all living birds) are known in basal dromaeosaurids, oviraptorosaurs, and scansoriopterygids (forming a fully fused sternum in some individuals of the first two clades), but are absent in troodontids and in the basal birds Archaeopteryx and Sapeornis (Clark et al., 1999; Norell and Makovicky, 1999; Wellnhofer and Tischlinger, 2004; Xu and Norell, 2004; Zhou and Zhang, 2003).
4.5. Propatagium
The lift generating effect of the propatagium must also be considered.
Richard E. Brown et al (1998)
Through flight experiments with live birds and computer modeling we define the aerodynamic contributions of the propatagium in avian flight. From flight trials we found that in House Sparrows, with all flight feathers removed except for the distal six primaries, the loss of approximately 50% of the propatagium's projected area and its cambered profile produced a significant reduction in the distance a bird was able to cover in flight. Removal of the secondary feathers, leaving six distal primaries and an intact propatagium, did not have a noticeable affect upon flight. From the computer model which is representative of the bird wing's mid-antebrachial chord (cambered propatagium, symmetrical musculoskeletal elements, and flat secondary flight feathers), we found that the propatagium: (1) produced the majority of the lift; (2) had a higher (relative to secondary feathers) production of lift in relation to its angle of attack, i.e., steeper lift-curve slope; and (3) produced more lift with a chord only 1/5 that of the feather subsection. We conclude that the cambered propatagium is the major lift generating component of the wing proximal to the wrist.
Cambered:
a. A slightly arched surface, as of a road, a ship's deck, an airfoil, or a snow ski.b. The condition of having an arched surface.
4.6. Semilunate carpal
Scansoriopterygids had a semilunate carpal (half-moon shaped wrist bone) that allowed for bird-like folding motion in the hand. This reduces drag on the upstroke which makes flapping flight possible, even without a derived supracoracoideus.
Mark N Putticket al (2014)
The origin of birds (Aves) is one of the great evolutionary transitions. Fossils show that many unique morphological features of modern birds, such as feathers, reduction in body size, and the semilunate carpal, long preceded the origin of clade Aves
The grouping of Oviraptorsauria plus Dromaeosauridae plus Troodontidae plus Avialae now has a name: Pennaraptora. A new detailed analysis of the evolution of the wrists of dinosaurs shows that the origin of the half-moon shaped wrist bone (semilunate carpal) is a pennaraptoran trait.
{kind=link}
XU Xing et al (2008)
With an estimated mass of 110 grams, Anchiornis is the smallest known non-avian theropod dinosaur. It exhibits some wrist features indicative of high mobility, presaging the wing-folding mechanisms seen in more derived birds and suggesting rapid evolution of the carpus.
4.7. Feathers
The relatively weak-looking flight feathers of basal birds [4], then, do not necessarily suggest that flight capability was poor, let alone entirely absent. Early birds and their close relatives could assemble an effective flying wing using multiple rows of relatively weak feathers, as demonstrated by Longrich's analysis of specimens of Archaeopteryx and Anchiornis [8].
4.8 Apomorphy of Avialae
Also, we can conclude that basal Euparaves were capable of flapping flight because their flight-related characteristics have often caused them to be placed within Avialae:
For example:
https://en.wikipedia.org/wiki/Pedopenna
https://en.wikipedia.org/wiki/Pedopenna
Pedopenna was originally classified as a paravian, the group of maniraptoran dinosaurs that includes both deinonychosaurs and avialans (the lineage including modern birds), but some scientists have classified it as a true avialan more closely related to modern birds than to deinonychosaurs.[3]
https://en.wikipedia.org/wiki/Avialae
Avialae is also occasionally defined as an apomorphy-based clade (that is, one based on physical characteristics). Jacques Gauthier, who named Avialae in 1986, re-defined it in 2001 as all dinosaurs that possessed feathered wings used in flapping flight, and the birds that descended from them.[8][9].
5.0 Oviraptors as secondarily flightless
Also see Appendix 3
TERESA MARYAŃSKA et al (2002)
http://agro.icm.edu.pl/agro/element/bwmeta1.element.agro-article-f38bf90a-7c6b-47fc-982a-30d0be44fd5e/c/app47-097.pdf
Oviraptorosauria is a clade of Cretaceous theropod dinosaurs of uncertain affinities within Maniraptoriformes. All previous phylogenetic analyses placed oviraptorosaurs outside a close relationship to birds (Avialae), recognizing Dromaeosauridae or Troodontidae, or a clade containing these two taxa (Deinonychosauria), as sister taxon to birds. Here we present the results of a phylogenetic analysis using 195 characters scored for four outgroup and 13 maniraptoriform (ingroup) terminal taxa, including new data on oviraptorids. This analysis places Oviraptorosauria within Avialae, in a sister−group relationship with Confuciusornis. Archaeopteryx, Therizinosauria, Dromaeosauridae, and Ornithomimosauria are successively more distant outgroups to the Confuciusornis−oviraptorosaur clade. Avimimus and Caudipteryx are successively more closely related to Oviraptoroidea, which contains the sister taxa Caenagnathidae and Oviraptoridae. Within Oviraptoridae, “Oviraptor” mongoliensis and Oviraptor philoceratops are successively more closely related to the
Conchoraptor−Ingenia clade. Oviraptorosaurs are hypothesized to be secondarily flightless. Emended phylogenetic definitions are provided for Oviraptoridae, Caenagnathidae, Oviraptoroidea, Oviraptorosauria, Avialae, Eumaniraptora, Maniraptora, and Maniraptoriformes.
Alan Feduccia et al (2015)
https://bio.unc.edu/files/2011/04/FeducciaCzerkas2015.pdf
Testing the neoflightless hypothesis: propatagium reveals flying ancestry of oviraptorosaurs
Testing the neoflightless hypothesis: propatagium reveals flying ancestry of oviraptorosaurs
We focus here on the discovery of an anatomical feature, the propatagium, which argues that Caudipteryx supports the neoflightless hypothesis—that is, it is derived from a flighted ancestry (Paul 2002)—and therefore its highly derived avian anatomy was selected for in an aerodynamic context.
The recognition of avian characters in oviraptorosaurs goes back to Elzanowski (1999, p. 311) who concluded that: ‘‘cranial similarities between oviraptorosaurs and ornithurine birds raise the possibility that oviraptorosaurs are the earliest known flightless birds.’’ We believe evidence now supports the view that many maniraptorans look avian, despite their inability to fly, because they were derived from basal volant birds and had become secondarily flightless (Fig. 1). Highly derived avian characteristics, such as the reduction of the manual digits in Caudipteryx, are so strikingly similar to that in extant birds that to conclude that this resemblance is only an exaptation defies the logical simplicity that it might look avian because it is a bird.
Considerable debate surrounds the numerousavian-like traits in core maniraptorans (oviraptorosaurs,troodontids, and dromaeosaurs), especially in theChinese Early Cretaceous oviraptorosaur Caudipteryx,which preserves modern avian pennaceous primary remigesattached to the manus, as is the case in modern birds.Was Caudipteryx derived from earth-bound theropod dinosaurs,which is the predominant view among palaeontologists,or was it secondarily flightless, with volant aviansor theropods as ancestors (the neoflightless hypothesis),which is another popular, but minority view. The discovery
here of an aerodynamic propatagium in several specimens
provides new evidence that Caudipteryx (and hence oviraptorosaurs)
represent secondarily derived flightless
ground dwellers, whether of theropod or avian affinity, and
that their presence and radiation during the Cretaceous mayhave been a factor in the apparent scarcity of many otherlarge flightless birds during that period.
Sorkin (2014)
Aerial ability in basal Deinonychosauria
The values of “potentially neoflightless character score and
grade” of 7.5-24 and 49-66 calculated by Paul(2002, table11.1)
for basal Protarchaeopteryx, Caudipteryx, Avimimus and derived
Oviraptoridae)members of Oviraptorosauria are second only
to those for Dromaeosauridae(38.25 and 83)among the non-avialan Theropoda. This makes it probable that the ancestors of the presently known flightless members of Oviraptorosauria possessed aerial ability superior to that of Anchiornis (“potentially
neoflightless character score& grade” for the more derived Troodontidae =17.5 and 45).
Altogether the evidence is substantive that Euparaves was a power-flying, primitive bird. It had numerous flying bird-like characteristics.
Oviraptors were secondarily flightless, having lost/reduced their flying bird-like characteristics as they adapted secondarily to life on the ground.
This hypothesis is supported by the evidence and does not require the extensive amount of exaptation and convergence that the current mainstream hypothesis requires. It is thus the more plausible, parsimonious hypothesis.
Notice the complexity of the wing and feather motion in the video below. It is implausible to claim that this complexity of feather and wing was pre-adapted and it is implausible to claim that it evolved multiple times independently (convergently).
https://en.wikipedia.org/wiki/Origin_of_avian_flight
The mechanics of an avian's wings involve a complex interworking of forces, particularly at the shoulder where most of the wings' motions take place. These functions depend on a precise balance of forces from the muscles, ligaments, and articular cartilages as well as inertial, gravitational, and aerodynamic loads on the wing.[2]
7.0 Future research
Altogether the evidence is substantive that Euparaves was a power-flying, primitive bird. It had numerous flying bird-like characteristics. And Oviraptors were secondarily flightless.
The next question is: What was the ancestor of flying Euparaves like? That is the direction future research could take.
=================================================================
SUPPLEMENTARY INFORMATION
APPENDIX 1
The evidence indicates that basal Euparaves were capable of flapping flight ("powered flight").
Hindwings provided additional lift
Vertebrate paleontologist Nick Longrich of the University of Bath in the United Kingdom, meanwhile, agrees that the large hindlimb feathers are an interesting discovery, but he suspects they may still have originally had an aerodynamic function. “I think they're airfoils,” he says—the structures on aircraft wings that provide lift. The hindlimb feathers “don't look like insulatory feathers to me: They're really long, they overlap the same way flight feathers do, and the fact that they curve is a characteristic flight feather feature.”https://www.researchgate.net/publication/236048162_Hind_Wings_in_Basal_Birds_and_the_Evolution_of_Leg_Feathers
Hind Wings in Basal Birds and the Evolution of Leg Feathers
Xiaoting Zheng,1,2* Zhonghe Zhou,3 Xiaoli Wang,1 Fucheng Zhang,3 Xiaomei Zhang,2 Yan Wang,1 Guangjin Wei,1 Shuo Wang,3,4 Xing Xu1,3* (2013)
Recent discoveries of large leg feathers in some theropods have implications for our understanding of the evolution of integumentary features on the avialan leg, and particularly of their relevance for the origin of avialan flight. Here we report 11 basal avialan specimens that will greatly improve our knowledge of leg integumentary features among early birds. In particular, they provide solid evidence for the existence of enlarged leg feathers on a variety of basal birds, suggest that extensively scaled feet might have appeared secondarily at an early stage in ornithuromorph evolution, and demonstrate a distal-to-proximal reduction pattern for leg feathers in avialan evolution.
The specimens described here collectively provide important new information about avialan hindlimb integumentary features, particularly in that they confirm the presence of a four-winged condition in basal birds. Large metatarsal feathers were first discovered in the basal dromaeosaurids Microraptor and Sinornithosaurus (1, 20–22), were subsequently reported in the enigmatic Pedopenna and the basal deinonychosaurians Anchiornis and Xiaotingia (2–4, 23), and can now also be definitively said to occur in the basal avialan Sapeornis. The morphology of the metatarsal feathers shows considerable variation among taxa known to possess these structures.
In Microraptor, the metatarsal feathers are proportionally large with highly asymmetrical vanes (1), whereas in Pedopenna, Anchiornis, and Sapeornis they are proportionally smaller and have nearly symmetrical vanes (2, 3). In all cases, however, the metatarsal feathers are similar in general arrangement (nearly perpendicular to the metatarsus, forming a large flat surface) and in having stiff vanes and curved rachises. These features suggest that the metatarsal feathers were aerodynamic in function(12), providing lift, creating drag, and/or enhancing maneuverability, and thus played a role in flight (1–7, 24, 25). The presence of metatarsal feathers with a probable aerodynamic function in both deinonychosaurians and avialans strongly supports the interpretation that leg feathers were an important factor in the origin of avialan flight, although the nature of their biomechanical contribution to flight ability in taxa that possessed them is debated (1–7, 24–26).
It would appear that the ability to take off from the ground began with Euavialae.
https://en.wikipedia.org/wiki/Euavialae
Anchiornis, Aurornis, Pedopenna, Xiaotingia and Zhongornis
These are all basal Euparaves. They could fly. They have been placed in different spots in different analyses by different researchers. We can conclude that they were capable of flapping flight because of their flight-related characteristics, which have often caused them to be placed within Avialae.
ANCHIORNIS
https://en.wikipedia.org/wiki/Aurornis

https://books.google.ca/books?id=V-okBwAAQBAJ&pg=PA344&lpg=PA344&dq=he+Rise+of+Birds:+225+Million+Years+of+Evolution,&source=bl&ots=wFx6eORFaR&sig=8E1j15GYFwJhINH1k5iRR9foHo8&hl=en&sa=X&ved=0ahUKEwiGoLnRnM_KAhVGgYMKHfTdAIEQ6AEIWzAO#v=onepage&q=Tetrapterygidae&f=false
The Rise of Birds: 225 Million Years of Evolution
Sankar Chatterjee
See Figure 3.16 page 45.
https://en.wikipedia.org/wiki/Scansoriopteryx

https://en.wikipedia.org/wiki/Euavialae
Euavialae (meaning "true winged birds") is a group of birds which includes all avialan species more closely related to modern birds, than to the primitive, long-tailed birds Archaeopteryx and Jeholornis.[1]
Anchiornis, Aurornis, Pedopenna, Xiaotingia and Zhongornis
These are all basal Euparaves. They could fly. They have been placed in different spots in different analyses by different researchers. We can conclude that they were capable of flapping flight because of their flight-related characteristics, which have often caused them to be placed within Avialae.
ANCHIORNIS
Anchiornis huxleyi was a small, paravian dinosaur with a triangular skull bearing several details in common with dromaeosaurids and troodontids.AURORNIS
Anchiornis is notable for its proportionally long forelimbs, which measured 80% of the total length of the hind limbs. This is similar to the condition in early avians such as Archaeopteryx, and the authors pointed out that long forelimbs are necessary for flight. Anchiornis also had a more avian wrist than other non-avialan theropods. The authors initially speculated that it would have been possible for Anchiornis to fly or glide. However, further finds showed that the wings of Anchiornis, while well-developed, were short when compared to later species like Microraptor, with relatively short primary feathers that had rounded, symmetrical tips, unlike the pointed, aerodynamically proportioned feathers of Microraptor.[2]
https://en.wikipedia.org/wiki/Aurornis
A phylogenetic analysis of Aurornis published in 2013 found that it belongs in the bird lineage, in a more basal position than Archaeopteryx.[2] The analysis was based on "almost 1,500 [anatomical] characteristics."[6] On the other hand, a phylogenetic analysis conducted by Brusatte et al. (2014) [see above] recovered Aurornis outside Avialae; it was recovered as a troodontid closely related to Anchiornis, Xiaotingia and Eosinopteryx.[7]
Pedopenna was originally classified as a paravian, the group of maniraptoran dinosaurs that includes both deinonychosaurs and avialans (the lineage including modern birds), but some scientists have classified it as a true avialan more closely related to modern birds than to deinonychosaurs.[3]
The bird-like characteristics of Pedopenna are further evidence of the dinosaur-bird evolutionary relationship. Apart from having a very bird-like skeletal structure in its legs, Pedopenna was remarkable due to the presence of long pennaceous feathers on the metatarsus (foot). Some deinonychosaurs are also known to have these 'hind wings', but those of Pedopenna differ from those of animals like Microraptor. Pedopenna hind wings were smaller and more rounded in shape. The longest feathers were slightly shorter than the metatarsus, at about 55 mm (2 in) long. Additionally, the feathers of Pedopenna were symmetrical, unlike the asymmetrical feathers of some deinonychosaurs and birds. Since asymmetrical feathers are typical of animals adapted to flying, it is likely that Pedopenna represents an early stage in the development of these structures. While many of the feather impressions in the fossil are weak, it is clear that each possessed a rachis and barbs, and while the exact number of foot feathers is uncertain, they are more numerous than in the hind-wings of Microraptor. Pedopenna also shows evidence of shorter feathers overlying the long foot feathers, evidence for the presence of coverts as seen in modern birds. Since the feathers show fewer aerodynamic adaptations than the similar hind wings of Microraptor, and appear to be less stiff, suggests that if they did have some kind of aerodynamic function, it was much weaker than in deinonychosaurs and birds. Xu and Zhang, in their 2005 description of Pedopenna, suggested that the feathers could be ornamental, or even vestigial. It is possible that a hind wing was present in the ancestors of deinonychosaurs and birds, and later lost in the bird lineage, with Pedopenna representing an intermediate stage where the hind wings are being reduced from a functional gliding apparatus to a display or insulatory function.[2]
The initial analysis by Xu et al. showed that Xiaotingia formed a clade with Archaeopteryx, Dromaeosauridae and Troodontidae to the exclusion of other groups traditionally seen as birds. Xu et al therefore (re)defined the concepts of Deinonychosauria and Avialae to the extent that Archaeopteryx and Xiaotingia belonged to the Deinonychosauria in the clade Archaeopterygidae.[1] This led to popular reports that "Archaeopteryx is no longer a bird",[2] although Xu et al noted that there are several competing definitions of the clade Aves currently in use, pointing out that their definitions are compatible with a traditional Aves with Archaeopteryx as a specifier.[1] This was challenged by an analysis using different methods published several months later however, in which Archaeopteryx was again recovered as an avialan, while Xiaotingia remained closely allied to Anchiornis within the Troodontidae.[3] In 2012, an expanded and revised version of the initial analysis also found Archaeopteryx to be avialan and Anchiornis to be troodontid, but recovered Xiaotingia as the most primitive member of the clade Dromaeosauridae rather than a close relative of Anchiornis within Troodontidae.[4]
ZHONGORNIS
https://en.wikipedia.org/wiki/Zhongornis
https://en.wikipedia.org/wiki/Zhongornis
Zhongornis (meaning "intermediate bird"[1]) is a genus of primitive birds that lived during the Early Cretaceous.
Reinterpretation of the Early Cretaceous maniraptoran (Dinosauria: Theropoda) Zhongornis haoae as a scansoriopterygid-like non-avian, and morphological resemblances between scansoriopterygids and basal oviraptorosaursJingmai K. O’CONNOR Corwin SULLIVAN
Revised anatomical interpretation of the tail and more detailed comparisons with non-avian dinosaurs strongly suggest that Zhongornis haoae is not a bird but a member or close relative of the enigmatic maniraptoran clade Scansoriopterygidae. Although the poor preservation and immature ontogenetic status of all known specimens of this clade prevent detailed comparisons, proportions of the hand support a close affinity with Zhongornis, while at the same time revealing significant differences between this genus and known basal birds. Zhongornis also bears some similarity to basal oviraptorosaurs, supporting the hypothesis that the Jurassic scansoriopterygids may be stem-group relatives of the Cretaceous Oviraptorosauria. This suggests that the Aves + Scansoriopterygidae clade, as resolved here including Zhongornis, may be an artifact of homoplasy and the currently limited information available for scansoriopterygids. Although we consider Zhongornis too poorly preserved and the current information too limited to definitively reassign this taxon to Scansoriopterygidae, we consider that there is ample evidence to strongly question the previous assignment of this taxon to Aves.
Tetrapterygidae (meaning "four-wings") is a group of four-winged dinosaurs proposed by Sankar Chatterjee in the second edition of his book The Rise of Birds: 225 Million Years of Evolution, where he included Microraptor, Xiaotingia, Aurornis, and Anchiornis.[1] The group was named after the characteristically long flight feathers on the legs of all included species, as well as the theory that the evolution of bird flight may have gone through a four-winged (or "tetrapteryx") stage, first proposed by naturalist William Beebe in 1915.[2] Chatterjee suggested that all dinosaurs with four wings formed a natural group exclusive of other paravians, and that this family was the sister taxon to the group Avialae, although most phylogenetic analyses have placed the animals of his Tetrapterygidae elsewhere in Paraves, such as Xiaotingia, Aurornis, and Anchiornis being placed in Avialae.[3]
https://books.google.ca/books?id=V-okBwAAQBAJ&pg=PA344&lpg=PA344&dq=he+Rise+of+Birds:+225+Million+Years+of+Evolution,&source=bl&ots=wFx6eORFaR&sig=8E1j15GYFwJhINH1k5iRR9foHo8&hl=en&sa=X&ved=0ahUKEwiGoLnRnM_KAhVGgYMKHfTdAIEQ6AEIWzAO#v=onepage&q=Tetrapterygidae&f=false
The Rise of Birds: 225 Million Years of Evolution
Sankar Chatterjee
See Figure 3.16 page 45.
Pascal Godefroit1 , Andrea Cau2 , Hu Dong-Yu3,4, François Escuillie´5 , Wu Wenhao6 & Gareth Dyke7 (2013a)
A Jurassic avialan dinosaur from China resolves the early phylogenetic history of birds
Epidendrosaurus, Epidexipteryx and Eosinopteryx, also from the Middle–Late Jurassic of northeastern China, are here regarded as basal, non-eumaniraptoran paravians. Thus our phylogeny is entirely consistent with the presence of a tetrapterygian condition (= four winged) and elongated rectrices in basal eumaniraptorans. We also postulate a single origin for typical forewing-powered flight, generally inferred to be present only in more derived birds5,24; shifting Archaeopteryx into deinonychosaurs3,4 minimally implies two origins (in Archaeopteryx and in ‘true’ birds) or a much more complex situation, with an earlier origin close to the base of Paraves for forewing-driven flight and subsequent modifications to the tetrapterygian condition in various deinonychosaurs5 . These relationships are also consistent with the recent discovery of potentially fourwinged flight surfaces in a range of Mesozoic basal birds25.
This new comprehensive phylogeny shows that basal avialans (Aurornis, Anchiornis, Xiaotingia) were already diversified in northern China during the Middle–Late Jurassic.
Scansoriopterygidae
We can conclude that Scansoriopterygids were capable of flapping flight because their flight-related characteristics have often caused them to be placed within Avialae.
Scansoriopteryx ("climbing wing") is a genus of avialan dinosaur. Described from only a single juvenile fossil specimen found in Liaoning, China, Scansoriopteryx is a sparrow-sized animal that shows adaptations in the foot indicating an arboreal (tree-dwelling) lifestyle. It possessed an unusual, elongated third finger. The type specimen of Scansoriopteryx also contains the fossilized impression of feathers.[1]
Scansoriopteryx lent its name to the family Scansoriopterygidae. Studies of dinosaur relationships have found Scansoriopteryx to be a close relative of true birds and a member of the clade Avialae.[11]
Scansoriopteryx heilmanni (and its likely synonym Epidendrosaurus ninchengensis) was the first non-avian dinosaur found that had clear adaptations to an arboreal or semi-arboreal lifestyle–it is likely that they spent much of their time in trees.
A monophyletic Scansoriopterygidae was recovered by Godefroit et al. (2013); the authors found scansoriopterygids to be basalmost members of Paraves and the sister group to the clade containing Avialae and Deinonychosauria.[10] Agnolín and Novas (2013) recovered scansoriopterygids as non-paravian maniraptorans andthe sister group to Oviraptorosauria.[11]
Both juvenile scansoriopterygid specimens preserve impressions of simple, down-like feathers, especially around the hand and arm. The longer feathers in this region led Czerkas and Yuan to speculate that adult scansoriopterygids may have had reasonably well-developed wing feathers which could have aided in leaping or rudimentary gliding, though they ruled out the possibility that Scansoriopteryx could have achieved powered flight. Like other maniraptorans, scansoriopterygids had a semilunate carpal (half-moon shaped wrist bone) that allowed for bird-like folding motion in the hand. Even if powered flight was not possible, this motion could have aided maneuverability in leaping from branch to branch.[4]
In a 2007 cladistic analysis of relationships among coelurosaurs, Phil Senter found Scansoriopteryx to be the closest dinosaurian relative of avian birds, and amember of the clade Avialae.[7] This view was supported by a second phylogenetic analysis performed by Zhang et al. in 2008.
The investigations – published in Journal of Ornithology – found a combination of plesiomorphic or ancestral non-dinosaurian traits along with highly derived unambiguous birdlike features. The researchers specifically note the primitive elongated feathers on the fore- and hind limbs, suggesting Scansoriopteryx is an ancestral form of early birds that had mastered basic aerodynamic manoeuvres of parachuting or gliding from trees.
These findings fulfil a prediction first made in the 1900s that the ancestors of birds didn’t evolve from dinosaurs, but instead from earlier arboreal archosaurs which originated flight according to the tree-down scenario. These small tree-dwelling archosaurs had improved ability to fly, with feathers that enabled them to at least glide. This ‘tree-down’ view is in contrast with the ‘ground-up’ view many palaeontologists side with.
Scansoriopteryx
APPENDIX 2
Regime shift at Paraves, rather than Aves
Before the origin of Aves, on the branch leading to Paraves, high rates of evolution led to a smaller body size and a relatively larger forelimb in Paraves. These changes are on a single branch leading to Paraves, representing a shift to a new smaller size and larger forelimb at this point.
Paraves, rather than Aves alone, shifted to a different evolutionary model relative to other coelurosaurian theropods (Table 2). On all trees and for both femur and forelimb size, the model with a regime shift at Paraves, rather than Aves, is favored (Table S10). We found strong support for a reduction in femur length within Paraves (57.5 mm for Paraves, 147.9 mm for other coelurosaurians) with weaker evidence for a concurrent reduction in evolutionary rates (Table 2). Importantly, the best-fitting paravian regime-shift model for femur length is substantially better than the best-fitting avian regime shift model (ΔAICc = 6.15; Table 1). The paravian forelimb reduced slightly in size (Paraves = 158.4 mm; other taxa = 234.4 mm; Table 2). In contrast, the equivalent model that incorporates a reduction in rate within Aves (Table 2) is an inferior fit (ΔAICc = 2.48).
Here again, the lineage leading to birds stands out as an exception, with maniraptoran theropods sustaining high rates of size evolution relative to other dinosaur lineages [12]. A third recent study [13] employing likelihood methods capable of detecting branch-specific rate shifts places the shift to higher rates of size evolution on the branch leading to Paraves (the clade uniting birds, dromaeosaurids, and troodontids).
A Basal Dromaeosaurid and Size Evolution Preceding Avian Flight
Fossil evidence for changes in dinosaurs near the lineage leading to birds and the origin of flight has been sparse. A dinosaur from Mongolia represents the basal divergence within Dromaeosauridae. The taxon's small body size and phylogenetic position imply that extreme miniaturization was ancestral for Paraves (the clade including Avialae, Troodontidae, and Dromaeosauridae), phylogenetically earlier than where flight evolution is strongly inferred.
This distal-first development led to a four-winged condition at the base of the Paraves.
Amy M. Balanoff, Gabe S. Bever, Timothy B. Rowe & Mark A. Norell (2013)
Features that were once considered exclusive to modern birds, such as feathers and a furcula, are now known to have first appeared in non-avian dinosaurs1 . However, relatively littleis known of the early evolutionary history of the hyperinflated brain that distinguishes birds from other living reptiles and provides the important neurological capablities required by flight2 . Here we use high-resolution computed tomography to estimate and compare cranial volumes of extant birds, the early avialan Archaeopteryx lithographica, and a number of non-avian maniraptoran dinosaurs that are phylogenetically close to the origins of both Avialae and avian flight. Previous work established that avian cerebral expansion began early in theropod history and that the cranial cavity of Archaeopteryx was volumetrically intermediate between these early forms and modern birds3,4. Our new data indicate that the relative size of the cranial cavity of Archaeopteryx is reflective of a more generalized maniraptoran volumetric signature and in several instances is actually smaller than that of other non-avian dinosaurs. Thus, bird-like encephalization indices evolved multiple times, supporting the conclusion that if Archaeopteryx had the neurological capabilities required of flight, so did at least some other non-avian maniraptorans. This is congruent with recent findings that avialans were not unique among maniraptorans in their ability to fly in some form5,6.
The brain of Archaeopteryx is not volumetrically avian but conforms to the expectation of a generalized paravian. Thus, if Archaeopteryx has a ‘flight-ready’ brain3 , which is almost certainly the case given its postcranial morphology, then so did other paravians. The hypothesis that dromaeosaurs and troodontids had the neurological capabilities required of powered flight, gliding, or some intermediate condition is congruent with the discovery of the ‘four-winged’ deinonychosaurs, Microraptor zhaoianus20 and Anchiornis huxleyi5,6
http://www.bris.ac.uk/news/2014/february/origin-of-birds.html
Mark Puttick and colleagues investigated the rates of evolution of the two key characteristics that preceded flight: body size and forelimb length. In order to fly, hulking meat-eating dinosaurs had to shrink in size and grow much longer arms to support their feathered wings.
"We were really surprised to discover that the key size shifts happened at the same time, at the origin of Paraves," said Mr Puttick of Bristol's School of Earth Sciences. "This was at least 20 million years before the first bird, the famous Archaeopteryx, and it shows that flight in birds arose through several evolutionary steps."
Roger B. J. Benson,1,* Nicolás E. Campione,2,3 Matthew T. Carrano,4 Philip D. Mannion,5Corwin Sullivan,6 Paul Upchurch,7 and David C. Evans3,8 (2014)
Rates of Dinosaur Body Mass Evolution Indicate 170 Million Years of Sustained Ecological Innovation on the Avian Stem Lineage
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4011683/
Rates of Dinosaur Body Mass Evolution Indicate 170 Million Years of Sustained Ecological Innovation on the Avian Stem Lineage
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4011683/
High rates were maintained only on the evolutionary line leading to birds, which continued to produce new ecological diversity not seen in other dinosaurs. Small body size might have been key to maintaining evolutionary potential (evolvability) in birds, which broke the lower body size limit of about 1 kg seen in other dinosaurs. Our results suggest that the maintenance of evolvability in only some lineages explains the unbalanced distribution of morphological and ecological diversity seen among groups of animals, both extinct and extant. Important living groups such as birds might therefore result from sustained, rapid evolutionary rates over timescales of hundreds of millions of years.
T. Alexander Dececchi, Hans CE Larsson, Michael B Habib (2016)
The wings before the bird: an evaluation of flapping-based locomotory hypotheses in bird antecedents
PREPRINT
https://peerj.com/preprints/1676.pdf
The glenoid fossa faces ventrolaterally in these taxa and only shifted to a more lateral configuration at Paraves (Makovicky and Zanno 2011; Turner et al. 2012)
Appendix 3
Oviraptors as secondarily-flightless
Here is an accumulation of material that supports the idea that Oviraptors were secondarily-flightless primitive birds. Being "secondarily-flightless" means they evolved from earlier flying primitive birds.
http://en.wikipedia.org/wiki/Oviraptorosauria
http://en.wikipedia.org/wiki/Oviraptorosauria
Oviraptorosaurs Temporal range: Cretaceous, 130–66 Ma
Oviraptorosaurs, like deinonychosaurs, are so bird-like that several scientists consider them to be true birds, more advanced than Archaeopteryx. Gregory S. Paul has written extensively on this possibility, and Teresa Maryańska and colleagues published a technical paper detailing this idea in 2002.[5][15][16] Michael Benton, in his widely-respected text Vertebrate Paleontology, also included oviraptorosaurs as an order within the class Aves.[17] However, a number of researchers have disagreed with this classification, retaining oviraptorosaurs as non-avialan maniraptorans slightly more primitive than the deinonychosaurs.[18]
Analyses like those of Maryanska et al (2002) and Osmólska et al. (2004) suggest that they may represent primitive flightless birds.[5][6]
The consensus view, based on several cladistic analyses, is that Caudipteryx is a basal (primitive) member of the Oviraptoridae, and the oviraptorids are nonavian theropod dinosaurs.[9] Incisivosaurus is the only oviraptorid that is more primitive.[10]
Halszka Osmólska et al. (2004) ran a cladistic analysis that came to a different conclusion. They found that the most birdlike features of oviraptorids actually place the whole clade within Aves itself, meaning that Caudipteryx is both an oviraptorid and a bird. In their analysis, birds evolved from more primitive theropods, and one lineage of birds became flightless, re-evolved some primitive features, and gave rise to the oviraptorids. This analysis was persuasive enough to be included in paleontological textbooks like Benton's Vertebrate Paleontology (2005).[11] The view that Caudipteryx was secondarily flightless is also preferred by Gregory S. Paul,[12] Lü et al.,[13] and Maryańska et al.[14]
Avialan status for Oviraptorosauria (2002)
TERESA MARYAŃSKA, HALSZKA OSMÓLSKA, and MIECZYSŁAW WOLSANOviraptorosauria is a clade of Cretaceous theropod dinosaurs of uncertain affinities within Maniraptoriformes. All previous phylogenetic analyses placed oviraptorosaurs outside a close relationship to birds (Avialae), recognizing Dromaeosauridae or Troodontidae, or a clade containing these two taxa (Deinonychosauria), as sister taxon to birds. Here we present the results of a phylogenetic analysis using 195 characters scored for four outgroup and 13 maniraptoriform (ingroup) terminal taxa, including new data on oviraptorids. This analysis places Oviraptorosauria within Avialae, in a sister−group relationship with Confuciusornis. Archaeopteryx, Therizinosauria, Dromaeosauridae, and Ornithomimosauria are successively more distant outgroups to the Confuciusornis−oviraptorosaur clade. Avimimus and Caudipteryx are successively more closely related to Oviraptoroidea, which contains the sister taxa Caenagnathidae and Oviraptoridae. Within Oviraptoridae, “Oviraptor” mongoliensis and Oviraptor philoceratops are successively more closely related to the
Conchoraptor−Ingenia clade. Oviraptorosaurs are hypothesized to be secondarily flightless. Emended phylogenetic definitions are provided for Oviraptoridae, Caenagnathidae, Oviraptoroidea, Oviraptorosauria, Avialae, Eumaniraptora, Maniraptora, and Maniraptoriformes.
Secondarily flightless birds or Cretaceous non-avian theropods?
Recent studies by Varricchio et al. reveal that males cared for the eggs of troodontids and oviraptorids, so-called "non-avian theropods" of the Cretaceous, just as do those of most Paleognathic birds (ratites and tinamous) today. Further, the clutches of both groups have large relative volumes, and consist of many eggs of relatively large size. By comparison, clutch care by most extant birds is biparental and the clutches are of small relative volume, and consist of but few small eggs. Varricchio et al. propose that troodontids and oviraptorids were pre-avian and that paternal egg care preceded the origin of birds. On the contrary, unmentioned by them is that abundant paleontological evidence has led several workers to conclude that troodontids and oviraptorids were secondary flightless birds. This evidence ranges from bird-like bodies and bone designs, adapted for climbing, perching, gliding, and ultimately flight, to relatively large, highly developed brains, poor sense of smell, and their feeding habits.
It is likely that some of these groups had both flying AND later secondarily-flightless members.
With the benefit of hindsight it is easy to see that if fossils of the small flying dromaeosaurs from China had only been discovered before the larger flightless dromaeosaurs like Deinonychus or Velociraptor were found, the interpretations of the past three decades on how birds are related to dinosaurs would have been significantly different. If it had already been established that dromaeosaurs were birds that could fly, then the most logical interpretation of larger flightless dromaeosaurs found afterwards would have to be that they represented birds, basically like the prehistoric equivalent of an Ostrich, which had lost their ability to fly.
Another point to consider is the following:
If the common ancestor (the basalmost Euparaves) was ground dwelling, then there have never been any secondarily-flightless primitive birds ever found:
Alan Feduccia1• Stephen A. Czerkas2, (2015)
Testing the neoflightless hypothesis: propatagium reveals flying ancestry of oviraptorosaurs
The absence in the Cretaceous fossil record of numerous large secondarily flightless birds has been considered a complex unsolved mystery (Feduccia 2012).http://www.bio.fsu.edu/James/Ornithological%20Monographs%202009.pdf
The loss of flight is of such common occurrence within Aves that it should be expected to have occurred any time after flight was initially achieved.
If some maniraptorans were birds, and if birds were not theropods, similarities between maniraptorans and theropods could be readily explained by convergence on a cursorial morphotype subsequent to the loss of flight. Even distantly related reptiles could converge closely, in some cases almost indistinguishably, on the theropod morphotype through the acquisition of cursoriality, as the case of Effigia, noted above, dramatically demonstrates (Nesbitt and Norell 2006, Nesbitt 2007).
Appendix 4
"Paraves" is a stem-based (branch-based) clade defined as:
"All dinosaurs which are more closely related to birds than to oviraptorosaurs."
and
"The first common ancestor of birds, dromaeosaurids, and troodontids which was not also ancestral to oviraptorosaurs."
Being "stem-based" means that "Paraves" includes not only the Paraves node but also the line preceding it, back to the previous node Pennaraptora (but not including Pennaraptora).
With the new understanding of Oviraptors as secondarily flightless primitive birds, the "Paraves" clade no longer has a meaning because there is no line preceding the Paraves node back to "Pennaraptora". So "Paraves loses its stem-based meaning.
Apomorphy of node based Euparaves:
"the clade stemming from the first panavian with ... remiges and rectrices, that is, enlarged, stiff-shafted, closed-vaned (= barbules bearing hooked distal pennulae), pennaceous feathers arising from the distal forelimbs and tail".[2]
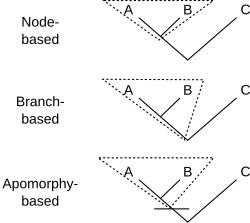
=============================================================
REFERENCES
Ingi Agnarssona,b* and Jeremy A. Millerc (2008)
Is ACCTRAN better than DELTRAN?
Unenlagiid theropods: are they members of the Dromaeosauridae (Theropoda, Maniraptora)?
Avian Ancestors
Linking the evolution of body shape and locomotor biomechanics in bird-line archosaurs
Evolutionary origins of the avian brain
Roger B. J. Benson,1,* Nicolás E. Campione,2,3 Matthew T. Carrano,4 Philip D. Mannion,5Corwin Sullivan,6 Paul Upchurch,7 and David C. Evans3,8 (2014)
Rates of Dinosaur Body Mass Evolution Indicate 170 Million Years of Sustained Ecological Innovation on the Avian Stem Lineage
Exploring macroevolution using modern and fossil data
Journal of Experimental Zoology
Gradual Assembly of Avian Body Plan Culminated in Rapid Rates of Evolution across the Dinosaur-Bird Transition
Luis M. Chiappe, Lawrence M. Witmer (2002)
Mesozoic Birds: Above the Heads of Dinosaurs
Czerkas, Yuan (2002)
A pre-Archaeopteryx troodontid theropod from China with long feathers on the metatarsus
Gregory M. Erickson,1,6,* Oliver W. M. Rauhut,2 Zhonghe Zhou,3 Alan H. Turner,4,6Brian D. Inouye,1 Dongyu Hu,5 and Mark A. Norell6 (2009)
Was Dinosaurian Physiology Inherited by Birds? Reconciling Slow Growth in Archaeopteryx
Feduccia, Czerkas (2014)
Jurassic archosaur is a non-dinosaurian bird
Alan Feduccia (1999)
The Origin and Evolution of Birds
Yale University Press
Alan Feduccia1• Stephen A. Czerkas2, (2015)
Testing the neoflightless hypothesis: propatagium reveals flying ancestry of oviraptorosaurs
Journal of Ornithology
Teresa J. Feo, Daniel J. Field, Richard O. Prum (2015)
Christian Foth1,2, Helmut Tischlinger3 & Oliver W. M. Rauhut1,2 (2014)
New specimen of Archaeopteryx provides insights
into the evolution of pennaceous feathers
Pascal Godefroit1 , Andrea Cau2 , Hu Dong-Yu3,4, François Escuillie´5 , Wu Wenhao6 & Gareth Dyke7(2013a)
A Jurassic avialan dinosaur from China resolves the early phylogenetic history of birdshttps://bioloskiblog.files.wordpress.com/2013/06/nature2013godefroit.pdf
Pascal Godefroit, Helena Demuynck, Gareth Dyke, Dongyu Hu, François Escuillié & Philippe Claeys (2013b)
Reduced plumage and flight ability of a new Jurassic paravian theropod from China
HINDWING FUNCTION IN FOUR-WINGED FEATHERED DINOSAURS
American Society of Biomechanics
Frances C. James1 and John A. Pourtless IV2 (2009)
Cladistics and the Origin of Birds: A Review and Two New Analyses
Daniel T. Ksepka (2014)
Evolution: A Rapid Flight towards Birds
Nicholas R. Longrich, Jakob Vinther, Qingjin Meng, Quangguo Li, Anthony P. Russell(2012)
Primitive Wing Feather Arrangement in Archaeopteryx lithographica andAnchiornis huxleyi
TERESA MARYAŃSKA, HALSZKA OSMÓLSKA, and MIECZYSŁAW WOLSAN (2002)
Avialan status for Oviraptorosauria
ACTA PALAEONTOLOGICA POLONICA
O'Connor and Sullivan, (2014)
Reinterpretation of the Early Cretaceous maniraptoran (Dinosauria: Theropoda) Zhongornis haoae as a scansoriopterygid-like non-avian, and morphological resemblances between scansoriopterygids and basal oviraptorosaurs
Institute of Vertebrate Paleontology and Paleoanthropology
STORRS L. OLSON; ALAN FEDUCCIA (1979)
Flight capability and the pectoral girdle of Archaeopteryx
https://repository.si.edu/bitstream/handle/10088/6524/VZ_93_Archaeopteryx.pdf?sequence=1&isAllowed=y
Mark N Puttick,1,2 Gavin H Thomas,3 Michael J Benton,1 and P David Polly (2014)
HIGH RATES OF EVOLUTION PRECEDED THE ORIGIN OF BIRDS
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4289940/
Corwin Sullivan, David W. E. Hone, Xing Xu, Fucheng Zhang (2010)
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4289940/
Sorkin (2014)
Aerial ability in basal Deinonychosauria
Corwin Sullivan, David W. E. Hone, Xing Xu, Fucheng Zhang (2010)
The asymmetry of the carpal joint and the evolution of wing folding in maniraptoran theropod dinosaurs
A Basal Dromaeosaurid and Size Evolution Preceding Avian Flight
Tim Wogan (2014)
Flight may have evolved multiple times in birds
Xiaoting Zheng,1,2* Zhonghe Zhou,3 Xiaoli Wang,1 Fucheng Zhang,3 Xiaomei Zhang,2 Yan Wang,1 Guangjin Wei,1 Shuo Wang,3,4 Xing Xu1,3* (2013)
Hind Wings in Basal Birds and the Evolution of Leg Feathers
A new maniraptoran dinosaur from China with long feathers on the metatarsus
http://link.springer.com/article/10.1007%2Fs00114-004-0604-y
http://www.ivpp.cas.cn/qt/papers/201403/P020140314318789141987.pdf
http://www.ivpp.cas.cn/qt/papers/201403/P020140314318789141987.pdf
XU Xing1† , ZHAO Qi1 , NORELL Mark2 , SULLIVAN Corwin1 , HONE David1 , ERICKSON Gregory2,3, WANG XiaoLin1 , HAN FengLu1,4 & GUO Yu1,4 (2008)
A new feathered maniraptoran dinosaur fossil that fills a morphological gap in avian origin
Xu et al, (2009)
A Jurassic ceratosaur from China helps clarify avian digital homologies
http://www.nature.com/articles/nature08124.epdf?referrer_access_token=1LIOYM249T2ALXmHhUVXQtRgN0jAjWel9jnR3ZoTv0NAxxXDTxDgb7tt7vNCs5i7CDx_p1E8pIL0dPMGIw0CIZ1LRnUZIDT1a3FIDY_UW4FRwpODRDVwWg-KbK448VK63yIXiGAa_H8fA42yVK8TsNhr_ASjWKKTbM-PJCMVzpKKElR4FEstewHl9DZGaHr9&tracking_referrer=www.nature.com
http://www.nature.com/articles/nature08124.epdf?referrer_access_token=1LIOYM249T2ALXmHhUVXQtRgN0jAjWel9jnR3ZoTv0NAxxXDTxDgb7tt7vNCs5i7CDx_p1E8pIL0dPMGIw0CIZ1LRnUZIDT1a3FIDY_UW4FRwpODRDVwWg-KbK448VK63yIXiGAa_H8fA42yVK8TsNhr_ASjWKKTbM-PJCMVzpKKElR4FEstewHl9DZGaHr9&tracking_referrer=www.nature.com
Nature
Xing Xu1,2, Hailu You3 , Kai Du4 & Fenglu Han2 (2011)
An Archaeopteryx-like theropod from China and the origin of Avialae
Xing Xu (2012)
Evolution: Taking Wing with Weak Feathers
Xing Xu,1 * Zhonghe Zhou,1 Robert Dudley,2 Susan Mackem,3 Cheng-Ming Chuong,4,7 Gregory M. Erickson,5 David J. Varricchio6 (2014)
An integrative approach to understanding bird origins
https://www.researchgate.net/publication/269715801_An_integrative_approach_to_understanding_bird_origins
A bizarre Jurassic maniraptoran from China with elongate ribbon-like feathers
Zhonghe Zhou (2014)
Current Biology
General
New insights into the origin of birds
------------------------------------------------------------
General Reference
The following study shows that there were 51 synapomorphies (unique defining characteristics) for Paraves (primitive birds). This means that of the 374 characteristics that were evaluated, 51 of them were different than the claimed dinosaur ancestor. This is more than 1 in 8.
Note that this number would be very much larger if the oviraptors etc were taken as secondarily flightless.
2011 study (Xu et al):
http://www.nature.com/nature/journal/v475/n7357/full/nature10288.html
http://www.ivpp.cas.cn/qt/papers/201403/P020140314389417822583.pdf
An Archaeopteryx-like theropod [Xiaotingia] from China and the origin of Avialae
Note that this number would be very much larger if the oviraptors etc were taken as secondarily flightless.
2011 study (Xu et al):
http://www.nature.com/nature/journal/v475/n7357/full/nature10288.html
http://www.ivpp.cas.cn/qt/papers/201403/P020140314389417822583.pdf
An Archaeopteryx-like theropod [Xiaotingia] from China and the origin of Avialae
Here we report a new Archaeopteryx-like theropod from China. This find further demonstrates that many features formerly regarded as being diagnostic of Avialae, including long and robust forelimbs, actually characterize the more inclusive group Paraves (composed of the avialans and the deinonychosaurs).
Paraves: 1.1, 10.1, 13.0, 14.0, 15.1, 20.1, 21.1, 28.1, 39.0, 61.1, 65.0, 66.0, 69.0, 79.0, 91.0,95.0, 96.1, 97.1, 106.0, 109.1, 119.1, 125.0, 127.1, 129.1, 137.1, 138.1, 139.1, 154.0, 155.1,156.1, 160.1, 166.0, 176.1, 179.1, 180.1, 184.1, 202.1, 221.1, 232.0, 237.1, 262.1, 267.1,277.2, 292.0, 304.2, 306.1, 319.1, 320.2, 336.1, 354.0, and 362.1
All pennaraptorans for which the integument is preserved (oviraptorosaurs, scansoriopterygids, dromaeosaurids, troodontids, avialians) show true broad pennaceous feathers on the arms and the tail. In eumaniraptorans (dromaeosaurids, troodontids, and basal avialians) long broad true feathers are present on the hindlimbs as well. Additionally, these same dinosaurs show some major transformations of the forelimb: elongate arms; semilunate carpal allowing for folding the hands; laterally oriented shoulder joints allowing the arms to stick out sideways; enlarged sternum for more powerful arm muscles. In modern birds these adaptations are useful in the flight stroke.
For reference:
Convergence on a cursorial (ground based) morphotype
Frances C. James et al (2009)
If some maniraptorans were birds, and if birds were not theropods, similarities between maniraptorans and theropods could be readily explained by convergence on a cursorial morphotype subsequent to the loss of flight. Even distantly related reptiles could converge closely, in some cases almost indistinguishably, on the theropod morphotype through the acquisition of cursoriality, as the case of Effigia, noted above, dramatically demonstrates (Nesbitt and Norell 2006, Nesbitt 2007).
Effigia is noted for its remarkable similarity to ornithomimid dinosaurs. Nesbitt, in 2007, showed that Effigiawas very similar to Shuvosaurus, and is definitely a member of the crurotarsan group Suchia (in the line leading towards modern crocodilians), and that its similarity to ornithomimids represents a case of "extreme" convergent evolution. Nesbitt also demonstrated that Shuvosaurus was the same animal as Chatterjeea, and that it belonged to an exclusive clade containing closely related suchians such as Shuvosaurus and Poposaurus (Poposauridae). Within this group, Effigia forms an even more exclusive clade with Shuvosaurus and the South American Sillosuchus(Shuvosaurinae).[1] In 2007, Lucas and others suggested "Effigia" was synonymous with "Shuvosaurus" and used the new combination "Shuvosaurus okeeffeae" for the animal.
An ossified sternum and uncinate processes are absent as in Anchiornis, Xiaotingia and troodontids. The humerus has a proximodistally long internal tuberosity, as in Anchiornis, Xiaotingia and other deinonychosaurs11
The length of manual phalanx IV-3 is considerably greater than the combined lengths of IV-1 and IV-2, a feature also seen in Anchiornis, Xiaotingia and other deinonychosaurs
Drawing:
http://csotonyi.com/Anchiornis_huxleyi_Csotonyi_wi.html
Xu et al 2014
Homologies
and homeotic transformation of the theropod 'semilunate' carpal.
Author information
Abstract
The homology of the 'semilunate' carpal, an important structure linking non-avian and avian dinosaurs, has been controversial. Here we describe the morphology of some theropod wrists, demonstrating that the 'semilunate' carpal is not formed by the same carpal elements in all theropods possessing this feature and that the involvement of the lateralmost distal carpal in forming the 'semilunate' carpal of birds is an inheritance from their non-avian theropod ancestors. Optimization of relevant morphological features indicates that these features evolved in an incremental way and the 'semilunate' structure underwent a lateral shift in position during theropod evolution, possibly as a result of selection for foldable wings in birds and their close theropod relatives. We propose that homeotic transformation was involved in the evolution of the 'semilunate' carpal. In combination with developmental data on avian wing digits, this suggests that homeosis played a significant role in theropod hand evolution in general.