This site presents the idea that birds developed from flying pterosaurs.
This is a credible alternative to the current, mainstream idea that birds developed from land-based dinosaurs.
'Beautiful' dinosaur tail found preserved in amber
The tail of a feathered dinosaur has been found perfectly preserved in amber from Myanmar.
The one-of-a-kind discovery helps put flesh on the bones of these extinct creatures, opening a new window on the biology of a group that dominated Earth for more than 160 million years.
The branched feathers have a weak pennaceous arrangement of barbs consistent with non-avialan coelurosaurs, particularly paravians. Although the feathers are somewhat pennaceous, none of the observed osteological features preclude a compsognathid [28] affinity. The presence of pennaceous feathers in pairs down the length of the tail may point toward a source within Pennaraptora [9], placing a lower limit on the specimen’s phylogenetic position. However, the distribution and shape of the feathers only strongly supports placement crownward of basal coelurosaurs, such as tyrannosaurids and compsognathids. In terms of an upper limit, the specimen can be confidently excluded from Pygostylia; in addition, it can likely be excluded from the long-tailed birds, based on pronounced ventral grooves on the vertebral centra.
That is a very nice Pennaraptoran/Paravian. It is not a dinosaur. The problem for the dinosaur to bird theory is that there is no connection between Pennaraptoran/Paravians and coelurosaur dinosaurs.
The plumulaceous (downy) and pennaceous feathers of Pennaraptora/Paraves evolved from Pterosaur pycnofibres and actinofibrils.
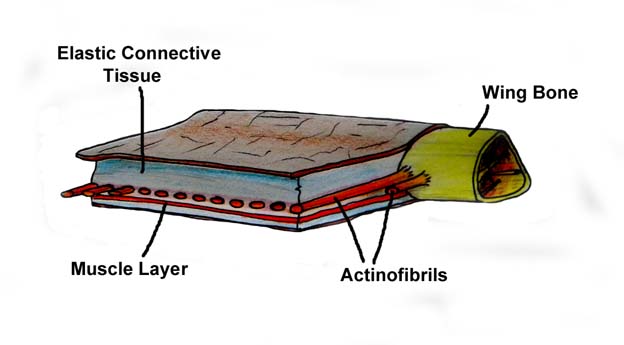
Pycnofibres covered the body, while the actinofibrils were covered by the wing membrane. Pycnofibres are comparable to Stage II feathers that have unfurled out of their sheaths. Actinofibrils are comparable to Stage IIIa feathers that are still within their sheaths.
While historically thought of as simple, leathery structures composed of skin, research has since shown that the wing membranes of pterosaurs were highly complex and dynamic structures suited to an active style of flight. The outer wings (from the tip to the elbow) were strengthened by closely spaced fibers called actinofibrils.[17] The actinofibrils themselves consisted of three distinct layers in the wing, forming a crisscross pattern when superimposed on one another.
The variation of space between adjacent actinofibrils in Jeholopterus, also reported in Rhamphorhynchus (Padian & Rayner 1993), suggests that those fibres were connected by some elastic tissue that enabled them to spread apart or join whenever necessary, making the actinopatagium more flexible (perhaps somewhat elastic
For the first time we observe actinofibrils (those fibers that support the wings) lying in multiple layers (not just a single one) and these tend to cross each other a little, though they are essentially subparallel. This tells us something about the structure and to a lesser extent function of the wing.
In this [Sordes], as in other pterosaurs, the wing fibers were embedded within the patagia [wing membranes] and typically measured a little less than one-tenth of a millimetre in diameter- about twice the thickness of a human hair. In some spots unravelled fibers reveal that they were composite structures composed of at least 20 or 30 very fine strands, wound together in a helical fashion. Each strand was only a few hundredth of a millimeter across and probably made of collagen a material that is common in the skin of vertebrates".
Wellnhofer [4, 5] and Padian [6, 7], following von Zittel [8],described a system of fine structural fibers investing the wing membrane,in a pattern similar to the orientation of the feather shafts of birds and the wing fingers of bats, both principal structural elements supporting the patagium and responsible for the transmission of aerodynamic force.
The wing membrane was supported and controlled through a system of stiffened, intercalated fibers,which were oriented like the main structural elements in the wings of birds and bats.
Actinofibrils are unusual structures and we are not sure exactly what they are composed of. The best guess is collagen, but it could also be cartilage or keratin. Determining this in fossils is obviously near impossible but all three are realistic possibilities, though of course collagen is the most likely given the position of the fibres inside the wing membrane (rather than on the surface) and they do not connect to the bones of the wing finger. They lie sub-parallel to the wing towards the wingtips and then sub-perpendicular as we move more proximally. There are no actinofibrils in the proximal wing close to the body, and they get more densely packed the further away you go.
Stage II — Origin of a collar with differentiated barb ridges results in a mature feather with a tuft of unbranched barbs and a basal calamus emerging from a superficial sheath.
Stage IIIa — Origin of helical displacement of barb ridges and the new barb locus results in a pinnate feather with an indeterminate number of unbranched barbs fused to a central rachis.
One distinctive feature of Scansoriopteryx is its elongated third finger, which is the longest on the hand, nearly twice as long as the second finger (in most theropod dinosaurs, the second finger is the longest). This is unlike the configuration seen in most other theropods, where the second finger is longest. The long wing feathers, or remiges, appear to attach to this long digit instead of the middle digit as in birds and other maniraptorans. Shorter feathers are preserved attached to the second finger.[6] A relative of Scansoriopteryx, Yi, suggests that this elongated third finger supported a membranous wingof some kind alongside feathers.[7]
The [rhamphorhynchoid] “hair-like” structures [pycnofibres]
are also unique in being preserved in fully three
dimensionally forms as compared to two
dimensional staining or impressions.The hairs [pycnofibres] are shown to be complex multi-strand structures instead of single strands or actual hairs.The complex nature of these filaments most closely resembles natal down feathers, but apparently without having barbules.As such, they may represent the earliest known form of feathers.This implies that such
integumentary structures may have originated
independently among pterosaurs from that of birds,
or that birds and pterosaurs may share a common
ancestor which had evolved this kind of insulation
before fight had been achieved in either group.
Feathers differ significantly from hair in that their multiple strands, the barbs,emanate from a single hollow structure, called the calamus. The integumentary structures seen in Pterorhynchus [a rhamphorhynchid]bear a striking similarity to that of a natal down feather with only the notable absence of having the additional barbules branching from the barbs. This absence is significant all the more because without the barbules, the barbs emanating from a calamus represents the hypothetical “Stage II” structurespeculated as being an incipient step in the evolution of feathers (Prum, 1999).
Proto-feathers have been attributed to two pterosaurs which are of similar animals (Ji and Yuan, 2002; Wang, et al., 2002). Even more so, the morphology details seen in Pterorhynchus demonstrate that the integumentary structures of pterosaurs are not like hair, but are analogous to being proto-feathers. Specifically, they resemble natal down feathers where individual filaments are seen to spread from a single follicle.
Therefore, the individual filaments are not representative of hair,
but are analogous to being the barbs of a feather.
Barbules, if present, cannot be discerned which
suggests that they either did not exist, or that the
limits of preservation have obscured them.
Nonetheless, the morphology of having several barbs stemming from a short calamus indicates that the body covering of Pterorhynchus are feather homologues. Without barbules, these structures would represent the second stage of feather
development as speculated by Prum (1999). The
feather homologues of Pterorhynchus also
demonstrate that a primary function achieved by
these plumulaceous feathers was that of thermal
insulation, and that feathers with a true rachis and
barbs aligned into well developed vanes represent
a derived condition.
The wing membranes are thought to have been stiffened by internal fibers, called aktinofibrils (Martill and Unwin, 1989; Wellnhofer, 1987, 1991). The distal end of a wing membrane is preserved in Pterorhychus which shows clear aktinofibrils that are aligned in parallel rows.
Kellner et al
On the tenopatagium close to the body and on the tail, a third type of fibre [pycnofibre] with somewhat diffuse edges is observed (figures 3a and and44a). Type C fibres can be easily separated from other fibres by their dark-brown colour and their general lack of organization. They are distributed along the body, the tail and the tip of the actinopatagium close to the fourth wing finger phalanx (figures 1, ,22 and and44c). Sometimes clustering together, they are not found covering the external portion of the plagiopatagium and are apparently rare on the actinopatagium.
As Wang et al. (2002) pointed out, these fibres are best interpreted as structures covering the body, commonly referred to as ‘hair’ or hair-like structures (e.g. Sharov 1971; Bakhurina & Unwin 1995). This pterosaur hair, which is not homologous to the mammalian hair (a protein filament that originates deep in the dermis and grows through the epidermis), is here called pycnofibre (from the Greek word pyknos, meaning dense, bushy). The pycnofibres are further formed by smaller fibrils of unknown nature. They were possibly mostly composed of keratin-like scales, feathers and mammalian hair.
Two other Chinese specimens were reported with integumental covering, coming from the same stratum (the Daohugou Bed) as Jeholopterus. So far we have not had the opportunity to examine this material. The first one is a small unnamed anurognathid with extensive preservation of soft tissue, including fibres that have been interpreted as protofeathers (Ji & Yuan 2002). The published pictures show that the soft tissue interpreted as protofeathers is of the same nature as the pycnofibres of Jeholopterus. There is no indication of branching structures that are expected for feather precursors.
Although no distinctive trailing edge is discernible, the wing membrane extends along the body and is connected to the hind limbs, reaching the ankle (Wang et al. 2002). While the distal portion of the plagiopatagium shows several layers of closely packed fibres (actinofibrils), the more proximal part lacks these structures. This confirms the observations of Schaller (1985), who recognized two distinct portions of the plagiopatagium, the actinopatagium and the tenopatagium, distinguished by the presence and absence of actinofibrils, respectively.
Compared with contour feathers, flight feathers have a larger pennaceous vane and a longer and thicker rachis. Wing flight feathers also have a longer calamus for insertion deeper into the follicle and anchor more securely to sustain its aerodynamic function
The bases of the flight feathers are covered with smaller contour feathers called coverts. There are several layers of coverts on the wing.
Both pycnofibres and actinofibrils are the same basic form. But they differ in that the pycnofibres proceeded through the stages to where the sheaths had disintegrated and the internal strands are visible on the surface. The actinofibrils did not proceed to that point. In the actinofibril, the internal strands (barbs) are still within the sheaths. And the sheaths are covered by the wing membrane.
The pycnofibres are on the body of the pterosaur. When the sheath disintegrates they unfurled into a form comparable to natal down and kept the body of the pterosaur warm.
The actinofibrils perform a different function. They stiffen and strengthen the wing membrane.
It is helpful to keep in mind that the actinofibrils and the membrane grew in synch as the pterosaur grew. (It is an example of facilitated variation.)
A schematic view of the three major structural components of the feather rachis. (a) (i) superficial layers of *fibres, the ultimate size-class in the hierarchy of feather keratin filaments (approx. 6 µm diameter), wound circumferentially round the rachis. (ii) The majority of the fibres extending parallel to the rachidial axis and through the depth of the cortex. Part of the section is peeled back to show why the fibres and even megafibrils are not usually recognized in histological sectioning, but rather only fibrils lower down the hierarchy (based on the electronic supplementary material, figure S2c). Any longitudinal section along the line of the arrows or at any point along the height of the fibre other than at the fibre surface (arrowheads) will fail to show the fibre. (iii) It shows the medulloid pith comprising gas-filled polyhedral structures (based on SEM images, electronic supplementary material, figures S5 and S6). Inset, part of a steel rebar with nodes, used in engineering technology to reinforce high-rise structures, analogous to rachidial fibres. (b) Schematic cross section of fibres and biodegraded ‘matrix’: (i) fibres; (ii) residual cytosol of keratinocytes presumably housing effete organelles and perhaps cytoskeletal elements—all degraded along with corneous envelope; (iii) interdigitating plasma membrane of the original keratinocytes with associated corneous envelope proteins. (c) A schematic three-dimensional cross section of the rachis showing approximate thickness (based on SEMs) of the three keratin layers comprising, (i) circumferential and (ii) longitudinal fibres of the cortex and (iii) polyhedra of medulloid pith. Asterisk denotes homologous with syncitial barbules. http://prumlab.yale.edu/sites/default/files/prum_n_brush_2002.pdf
Figure 2. Schematic Diagram of Helical Growth of Barb Ridges of a Pennaceous Feather The branched structure of the barbs and the rachis of a feather form by helical growth and fusion of barb ridges within the tubular feather germ. Feathers grow from the base. Barb ridges form at the new barb locus on the posterior midline of the collar and grow helically around the collar toward the anterior midline where they fuse to form the rachis ridge. Subsequent barb ridges fuse to the rachis ridge. In feathers with an afterfeather, the new barb locus divides into two laterally displaced new barb loci. Subsequently, new barb ridges grow helically both anteriorly to the main rachis and posteriorly to form the hyporachis and vane of the afterfeather. The main vane and the afterfeather form separate vanes united within a single feather by the calamus (Figure 1A). Pennaceous feathers obtain their planar form only after emerging from the cylindrical feather sheath when growth is complete. The obverse (upper) and reverse (lower) surfaces of the vane develop from the outer and inner surfaces of the cylindrical feather germ. Illustration based on Lucas and Stettenheim (1972).
See how it's covered in skin already? There's a tendon running between the shoulder and the wrist, just like in pterosaurs, that anchors a skin membrane called the propatagium. The ulna is covered in thick skin that anchors the flight feathers. In many birds, the bottom of the upper arm is loosely connected to the body by skin as well. So you can imagine these membranes becoming larger and more parachute-like.
Czerkas & Yuan also noticed tissue impressions coming off the ulna and third finger of Scansoriopteryx—those might be better interpreted (now) as a flight membrane.
There are indications from where the feathers emanate
below the ulna which suggest that a short patagium may
have been present. Unlike most of the wing feathers, there
appears to be a series of feathers that do not reach the bone
itself.
In barb medullary cells feather keratin is accumulated in few peripheral bundles that merge with those of cortical cells to form the wall of the ramus. The latter is joined with branching barbules.
What’s been mostly overlooked in discussions of Yi qi is that pennaraptoran maniraptorans already have patagia.
Look at the (perhaps familiar) pictures of nightjar wings below and observe all the ‘webbing’ that surrounds the fingers and arm. A membrane called the propatagium spans the space between the wrist and shoulder, and membranes run along the trailing (or posterior, or postaxial) edges of the hand and ulna too.
Xu et al. (2015) also note that these [Yi qi] patches have an unusual wrinkled texture, not typical of skin that would have been covered in filaments or feathers. But maybe they’re naked and wrinkled for taphonomic reasons.
As a Tet Zoo regular, you'll recognise these images from p. 19 of Katrina van Grouw's The Unfeathered Bird. They show the left wing of a European nightjar (Caprimulgus europaeus) in (at top) dorsal and ventral views. Note all the skin membranes around the more muscular parts of the wing. Image by Katrina van Grouw, used with permission.
The four patagia of the wing include the propatagium, where the wing and the neck join the thorax; the postpatagium, which is located at the caudal angle of the carpus; the metapatagium at the caudal junction of the thorax and the wing, and the alular patagium between the alula and the carpometacarpus.
Remiges (from the Latin for "oarsman") are located on the posterior side of the wing. Ligaments attach the long calami (quills) firmly to the wing bones, and a thick, strong band of tendinous tissue known as the postpatagium helps to hold and support the remiges in place.[2]
On the tenopatagium close to the body and on the tail, a third type of fibre with somewhat diffuse edges is observed (figures 3a and 4a). Type C fibres can be easily separated from other fibres by their dark-brown colour and their general lack of organization. They are distributed along the body, the tail and the tip of the actinopatagium close to the fourth wing finger phalanx (figures 1, 2 and 4c). Sometimes clustering together, they are not found covering the external portion of the plagiopatagium and are apparently rare on the actinopatagium.
Generally thicker than the actinofibrils (figure 4a), type C fibres have an average thickness ranging between 0.2 and 0.5 mm. In several places, it is clear that they are formed by smaller fibrils, the nature of which is unknown. The sediment between type C fibres tends to be light brown in colour, making the distinction of individual fibres more difficult. Several cross each other but lack the reticular pattern formed by the multi-layered superposition of the actinofibrils. In several areas, type C fibres are preserved associated with an amorphous whitish matter that has been interpreted as patches of the epidermis (and dermis, described earlier). Although not parallel to each other and lacking the organization of the actinofibrils, fibres C in most parts are generally displaced away from the skeleton.
As Wang et al. (2002) pointed out, these fibres are best interpreted as structures covering the body, commonly referred to as ‘hair’ or hair-like structures (e.g. Sharov 1971; Bakhurina & Unwin 1995). This pterosaur hair, which is not homologous to the mammalian hair (a protein filament that originates deep in the dermis and grows through the epidermis), is here called pycnofibre (from the Greek word pyknos, meaning dense, bushy). The pycnofibres are further formed by smaller fibrils of unknown nature. They were possibly mostly composed of keratin, like scales, feathers and mammalian hair.
One detail, however, of
feather development appears to violate von Baer’s
rule. During the development of the first feather
papillae in the embryo (before day 12 in the chick,
Gallus gallus), the barb ridge primordia appear
as longitudinal condensations within the feather
papillae before the follicle and collar are fully
formed (Lucas and Stettenheim, ’72). However, this
developmental event—the origin of the feather before
the follicle and collar—is clearly derived because
barb ridges would be unable to grow without
the spatial organization provided by the collar.
Dinosaurs did not have feathers, they had bristles. Dinosaur to bird proponents misclassify dinosaur bristles as (proto)feathers. There is no connection between the bristles in Coelurosaur dinosaurs and the pennaceous feathers in Pennaraptora/Paraves.
Morphotype 1 is known from the heterodontosaurid Tianyulong and the ceratopsian Psittacosaurus (Mayr et al., 2002; Zheng et al., 2009). Its main characteristic is being monofilament and relatively great length and rigidity. A variant of this morphotype is seen in Beipiaosaurus, which differs from those of Tianyulong and Psittacosaurus in its relatively great width ( Xu et al., 2009b). Morphotype 2 is a compound structure composed of multiple filaments joined basally. It is clearly present in Sinornithosaurus and Anchiornis, and probably also in Sinosauropteryx and Beipiaosaurus. Morphotype 3 is a distally branched filament, which is seen in the holotype of Sinornithosaurus millenii and probably in Beipiaosaurus (Xu et al., 1999). The main characteristic of this morphotype of feather is its barbs breaking off from the tip of a central filament and distally positioned short barbs. Morphotype 4 is a compound structure consisting of multiple filaments branching laterally from most of the length of a central filament. It is known in Sinornithosaurus, Anchiornis, Caudipteryx, Protarchaeopteryx, and probably Dilong as well (Xu et al., 2004). Morphotype 5 is only known in Epidexipteryx. It consists of parallel barbs arising from the edge of a membrane structure (Zhang et al., 2008b). Given its so unusual morphology, possibility of its being part of a more complete integumentary structure could not be completely excluded, particularly in consideration that morphotypes 2 and 4 display distally parallel barbs in some cases.
Among these defining features, tubular nature and filamentous morphology represent the earliest ones appearing in feather evolution and mark the origin of feathers as indicated by both paleontological and neontological data (Harris et al., 2002; Xu et al., 2009b). Feathers are thus here defined as integumentary structures that are tubular and filamentous in morphology. Follicle, hierarchical branches, and planar form are inferred to have evolved later in feather evolution.
Five major morphogenesis events are inferred to have occurred sequentially in feather evolution before the origin of the Aves and they are: 1) appearance of filamentous and tubular morphology, 2) formation of follicle and barb ridges, 3) appearance of rachis, 4) appearance of
planar form, and 5) formation of pennaceous barbules.
A notable feature is that the filaments in feather morphotype 2 are somewhat straplike, a feature also characteristic of barbs in modern feathers, yet the filaments in feather morphotype 2 are apparently proportionally wider than barbs in modern feathers. Recent developmental studies demonstrate the impossibility of separate formation of barb and barbule cells and
suggest that primitive feathers with only barbs but not barbules are unlikely to exist (Alibardi,
2005). If this holds true, some sort of simple, small barbules might be present in feather morphotype 2 or other primitive feathers.
A sudden appearance of a whole set of unique, complex developmental mechanisms and associated morphologies is also unlikely from the perspective of adaptation.
Morphotype 1 filaments are bristles. Morphotype 2 filaments are clustered bristles. "Morphotype 2" filaments are NOT formed from a follicle. They are not "joined basally". =================================================
Over the course of the last two decades, the understanding of the early evolution of feathers in nonavian dinosaurs has been revolutionized. It is now recognized that early feathers had a simple form comparable in general structure to the hairs of mammals. Insight into the prevalence of simple feathers throughout the dinosaur family tree has gradually arisen in tandem with the growing evidence for endothermic dinosaur metabolisms. This has led to the generally accepted opinion that the early feather coats of dinosaurs functioned as thermo insulation. However, thermo insulation is often erroneously stated to be a likely functional explanation for the origin of feathers. The problem with this explanation is that, like mammalian hair, simple feathers could serve as insulation only when present in sufficiently high concentrations. The theory therefore necessitates the origination of feathers en masse. We advocate for a novel origin theory of feathers as bristles.Bristlesare facial feathers common among modern birds that function like mammalian tactile whiskers, and are frequently simple and hair-like in form. Bristles serve their role in low concentrations, and therefore offer a feasible first stage in feather evolution.
http://www.ncbi.nlm.nih.gov/pubmed/12955841 Origin of archosaurian integumentary appendages: the bristles of the wild turkey beard express feather-type beta keratins.
Sawyer RH1, Washington LD, Salvatore BA, Glenn TC, Knapp LW.
The discovery that structurally unique "filamentous integumentary appendages" are associated with several different non-avian dinosaurs continues to stimulate the development of models to explain the evolutionary origin of feathers. Taking the phylogenetic relationships of the non-avian dinosaurs into consideration, some models propose that the "filamentous integumentary appendages" represent intermediate stages in the sequential evolution of feathers. Here we present observations on a unique integumentary structure, the bristle of the wild turkey beard, and suggest that this non-feather appendage provides another explanation for some of the "filamentous integumentary appendages." Unlike feathers, beard bristles grow continuously from finger-like outgrows of the integument lacking follicles. We find that these beard bristles, which show simple branching, are hollow, distally, and express the feather-type beta keratins. The significance of these observations to explanations for the evolution of archosaurian integumentary appendages is discussed.
Strangely, bristles somewhat like porcupine quills may have spread across most of the body of Pegomastax. Such bristles first appeared in a relative named Tianyulong recently discovered in China. Buried in lake sediments and covered by volcanic ash, Tianyulong was preserved with hundreds of bristles covering its body from its neck to the tip of its tail.
The feather-like structures found on the new heterodontosaur fossil [Tianyulong] are rigid, tubular and not that downy, You writes. They somewhat resemble relatively long, stiff, quills or bristles that have been reported on psittacosaurs — only the psittacosaur's are stiffer and more widely separated.
what we see in this heterodontosaur [Tianyulong] might be a separate evolution of some sort of projecting epidermal filament.
The filamentous integumentary structures in Jehol theropods
have been interpreted as protofeathers22. The presence of the similar
structures in IVPP V11579 [Dilong] provides the first direct evidence showing
that tyrannosauroids possessed protofeathers. Furthermore, the
filamentous protofeathers are branched as in other coelurosaurians22.
"Feathers, however, are hierarchically complex assemblages of numerous
evolutionary novelties—the feather follicle, tubular feather germ, feather branched structure,
interacting differentiated barbules—that have no homolog in any antecedent structures"
Initial
Development of a Feather Follicle
(A) Development of the epidermal feather placode
and dermal condensation. (B) Development of a short
bud or feather papilla. (C) Formation of the feather
follicle through the invagination of a cylinder of epidermal
tissue around the base of the feather papilla.
(D) Cross section of the feather follicle through the
horizontal plane indicated by the dotted line in C. The
invaginated tubular feather follicle is characterized by
a series of tissue layers (from peripheral to central):
the dermis of the follicle, the epidermis of the follicle
(outer epidermal layer), the follicle cavity or lumen
(the space between epidermal layers), the follicle collar
(inner epidermal layer or ramogenic zone), and
the dermal pulp (tissue at the center of the follicle).
The tubular feather germ grows by proliferation and
differentiation of keratinocytes in the follicle collar.
Summarized from Lucas and Stettenheim (1972) and
Prum (1999).
Developmental Model of the Origin and Diversification of Feather Follicles
An hypothesized transition series of evolutionary novelties in feather development, depicted as a series of
cross sections of the follicle collar—the innermost layer of epidermal tissue in the feather follicle that generates
or develops into the feather (Figure 3D) from Prum (1999). The model is based entirely on the hierarchical
details of feather development, and is independent of functional or phylogenetic assumptions. Each diagram
is oriented with the anterior surface of the follicle collar upward. The developmental novelties are labeled in
the stages at which they originate. Stage I—Origin of the undifferentiated collar through a cylindrical epidermal
invagination around the base of the feather papilla. Stage II—Origin of the differentiation of the inner layer
of the collar into longitudinal barb ridges and the peripheral layer of the collar into the feather sheath. Stage
III—Either of these developmental novelties could have occurred first, but both are required before Stage IV.
Stage IIIa—Origin of helical displacement of barb ridges and the new barb locus. Stage IIIb—Origin of paired
barbules from peripheral barb plates within the barb ridges. Stages IIIaIIIb—Origin of follicle capable of
both helical displacement and barbule plate differentiation. Stage IV—Origin of differentiated distal and proximal
barbules within barbule plates of barb ridges. Stage Va—Origin of lateral displacement of the new barb
ridge locus. Stage Vb—Origin of the division of posterior new barb locus into a pair of laterally displaced loci,
and opposing anterior and posterior helical displacement of barb ridges toward the main feather and afterfeather
of the follicle. See Prum (1999) for details of additional stages in the evolution of feather diversity
(Stages Vc–f).
Developmental Model of the Origin and Diversification of Feathers
A predicted transition series of feather follicles based on the hypothesized series of evolutionary novelties in
feather developmental mechanisms (Figure 4) from Prum (1999). Stage I—Origin of an undifferentiated tubular
collar yields the first feather, a hollow cylinder. Stage II—Origin of a collar with differentiated barb ridges
results in a mature feather with a tuft of unbranched barbs and a basal calamus emerging from a superficial
sheath. Stage IIIa—Origin of helical displacement of barb ridges and the new barb locus results in a pinnate
feather with an indeterminate number of unbranched barbs fused to a central rachis. Stage IIIb—Origin of
peripheral barbule plates within barb ridges yields a feather with numerous branched barbs attached to a basal
calamus. Stages IIIaIIIb—Origin of a feather with both a rachis and barbs with barbules creates a bipinnate,
open pennaceous structure. Stage IV—Origin of differentiated proximal and distal barbules creates the first
closed pennaceous vane. Distal barbules grew terminally hooked pennulae to attach to the simpler, grooved
proximal barbules of the adjacent barb (Figure 1B). Stage Va—Lateral displacement of the new barb locus
leads to the growth of a closed pennaceous feather with an asymmetrical vane resembling modern rectrices
and remiges. Stage Vb—Division and lateral displacement of the new barb loci yields opposing, anteriorly and
posteriorly oriented patterns of helical displacement, producing a main feather and an afterfeather with a
single calamus. The afterfeather could have evolved at any time following Stage IIIb, but likely occurred after
Stage IV based on modern aftershaft morphology. See Prum (1999) for details of additional stages in the
evolution of feather diversity.
By focusing on the evolution of the mechanisms
of feather development, Prum (1999)
proposed a detailed, testable model of the
evolutionary origin of feathers that is independent
of functional or phylogenetic
assumptions. The model proposed a five stage
transition series in the history of
feather diversity as a hypothesized sequence
of novelties in feather development (Figure
4). The model hypothesizes that the first
feather (Stage I) originated with the first
feather follicle—the cylindrical epidermal
invagination around the initial feather papilla.
Subsequent feather diversity evolved through
a series of derived developmental novelties
within the tubular intermediate epidermal layer of the follicle, called the follicle collar,
which generates the tubular feather germ.
After the origin of the follicle came the differentiation
of the follicle collar into barb ridges
that generate the barbs (Stage II). The model
proposes two alternative stages next—the origin of helical growth (Stage IIIa), or the origin
of barbule plate differentiation (Stage IIIb).
The model cannot differentiate between the
two alternative orders for these events (i.e.,
IIIa before IIIb, or IIIb before IIIa), but following
the evolution of both of these developmental
novelties came the capacity to grow
both kinds of branched structure typical of
modern feathers (Stage IIIab). The origin
of differentiated distal and proximal barbule
plates followed next (Stage IV). Finally, additional
developmental mechanisms evolved
and created further diversity in feather structure
(Stage Va–f).
Prum and Brush ignore the first two steps and label the C stage (seen above) as Stage I of their feather development stages. When researchers claim that dinosaurs have Stage I feathers, the evidence is that they had Stage B above and not Stage C. And Stage B is just a bristle. Note: When Prum and Brush showed that feathers did not evolve from scales, then the dinosaur to bird theorists were stuck. Pennaceous feathers did not evolve from scales nor from any dinosaur filament! http://people.eku.edu/ritchisong/feather_evolution.htm
Because birds evolved from reptiles and the integument of present-day reptiles (and most extinct reptiles including most dinosaurs) is characterized by scales, early hypotheses concerning the evolution of feathers began with the assumption that feathers developed from scales, with scales elongating, then growing fringed edges and, ultimately, producing hooked and grooved barbules (Figure 6 below). The problem with that scenario is that scales are basically flat folds of the integument whereas feathers are tubular structures. A pennaceous feather becomes ‘flat’ only after emerging from a cylindrical sheath (Prum and Brush 2002). In addition, the type and distribution of protein (keratin) in feathers and scales differ (Sawyer et al. 2000). The only feature shared by feathers and scales is that they both begin development as a morphologically distinct placode – an epidermal thickening above a condensation, or congregation, of dermal cells (see Figure 8 below). Feathers, then, are not derived from scales, but, rather, are evolutionary novelties with numerous unique features, including the feather follicle, tubular feather germ (an elevated area of epidermal cells), and a complex branching structure (Prum and Brush 2002
The origin of feathers is a specific instance of the much more
general question of the origin of evolutionary novelties—structures
that have no clear antecedents in ancestral animals and no
clear related structures (homologues) in contemporary relatives.
Although evolutionary theory provides a robust explanation
for the appearance of minor variations in the size and shape of
creatures and their component parts, it does not yet give as
much guidance for understanding the emergence of entirely
new structures, including digits, limbs, eyes and feathers.
This suggests that large pennaceous feathers first evolved distally on the hindlimbs, as on the forelimbs and tail. This distal-first development led to a four-winged condition at the base of the Paraves. (Hu et al 2009)
The significant lengthening and thickening of the forelimbs indicates a dramatic shift
in forelimb function at the base of the Paraves, which might be related to the appearance of a degree of aerodynamic capability. This hypothesis is consistent with the presence of flight feathers with asymmetrical vanes in both basal avialans and basal deinonychosaurs6,23.
Many dinosaurian groups, such as most ornithischians, the sauropodomorphs and the basal theropods, are not included in this simplified dinosaurian cladogram. The available specimens suggest that members of these groups had scaly skin, but the possibility that they are partially covered by filamentous integumentary structures cannot be completely excluded. Preservational factors make it difficult to observe the detailed structure of the filamentous feathers in available specimens of compsognathids, tyrannosauroids, and therizinosauroids, so a ‘?’ is used to indicate uncertainty regarding the presence of morphotypes 1, 3, 4 and 5 in these groups. On the basis of the anatomical, ontogenetic, and phylogenetic distribution patterns of known feather morphotypes among non-avian dinosaurs and early birds, morphotypes 1, 2 and 7 are inferred to have been lost in feather evolution, along with their associated developmental mechanisms.
Pennaceous feathers thus represented an exaptation and were later, in several lineages and following different patterns, recruited for aerodynamic functions. This indicates that the origin of flight in avialans was more complex than previously thought and might have involved several convergent achievements of aerial abilities.
For as long as dinosaurs have been known to exist, there has been speculation about their appearance. Fossil feathers can preserve the morphology of color-imparting melanosomes, which allow color patterns in feathered dinosaurs to be reconstructed. Here, we have mapped feather color patterns in a Late Jurassic basal paravian [Anchiornis] theropod dinosaur. Quantitative comparisons with melanosome shape and density in extant feathers indicate that the body was gray and dark and the face had rufous speckles. The crown was rufous, and the long limb feathers were white with distal black spangles. The evolution of melanin-based within-feather pigmentation patterns may coincide with that of elongate pennaceous feathers in the common ancestor of Maniraptora, before active powered flight. Feathers may thus have played a role in sexual selection or other communication.
Among extinct dinosaurs, feathers or feather-like integument have been discovered on dozens of genera via both direct and indirect fossil evidence. The vast majority of feather discoveries have been for coelurosaurian theropods. However, integument has also been discovered on at least three ornithischians, raising the likelihood that proto-feathers were also present in earlier dinosaurs, and perhaps even a more ancestral animal, in light of the pycnofibers of pterosaurs.
Note that the filamentous structures in some ornithischian dinosaurs (Psittacosaurus, Tianyulong and Kulindadromeus) and the pycnofibres found in some pterosaursmay or may not be homologous with the feathers of theropods.[45][62]
While it has been known since 2004, upon the description of Dilong, that at least some tyrannosauroids possessed filamentous "stage 1" feathers,[4] according to the feather typology of Richard Prum, Y. huali is currently the largest known species of dinosaur with direct evidence of feathers, forty times heavier than the previous record holder, Beipiaosaurus.[2][5] The feathers were long, up to 20 centimetres (7.9 in), and filamentous. Because the quality of the preservation was low, it could not be established whether the filaments were simple or compound, broad or narrow. The feathers covered various parts of the body. With the holotype they were present on the pelvis and the foot. Specimen ZCDM V5000 had feathers on the tail pointing backwards under an angle of 30° with the tail axis. The smallest specimen showed 20 centimetre (7.9 inch)-long filaments on the neck and 16 centimetre (6.3 inch)-long feathers at the upper arm.[2]
Feduccia, A.; Lingham-Soliar, T.; Hinchliffe, J. R. (2005). "Do feathered dinosaurs exist? Testing the hypothesis on morphological and paleontological evidence". Journal of Morphology. 266 (2): 125–166
Our findings show no evidence for the existence of protofeathers and consequently no evidence in support of the follicular theory of the morphogenesis of the feather. Rather, based on histological studies of the integument of modern reptiles, which show complex patterns of the collagen fibers of the dermis, we conclude that “protofeathers” are probably the remains ofcollagenous fiber “meshworks” that reinforced the dinosaur integument. These “meshworks” of the skin frequently formed aberrant patterns resembling feathers as a consequence of decomposition.
Note that Dilong was NOT Morphotype 4. It was NOT a "compound structure consisting of multiple filaments branching laterally from most of the length of a central filament."
Note that the filamentous structures in some ornithischian dinosaurs (Psittacosaurus, Tianyulong and Kulindadromeus) and the pycnofibres found in somepterosaurs may or may not be homologous with the feathers of theropods.[45][62]
Most of these are paravians with feathers. The others are non-paravian dinosaurs (ie actual dinosaurs). They have bristles orthe remains ofcollagenous fiber “meshworks”. There are no feathered dinosaurs.
It's possible that a few ornithischians, like those in the two photos above, separately evolved some kind of bristle for their own reasons, and that these bristles have no relation to the protofeathers of early theropods. It's also possible that the bristles on the above dinosaurs are homologous with theropod proto-feathers, and that the first dinosaurs all had some kind of fuzzy/bristly growths that were then later lost in most of the sauropods/ornithischians . . . or that the fuzz was reserved for baby dinosaurs, and only later spread to adults (although that doesn't explain the lack of feathers on the in-egg sauropod embryos.)
The ramifications of the
claims may best be understood in Prum and Brush’s
(2003, p. 92) own words: "The heterogeneity of the
feathers found on these dinosaurs is striking and
provides strong direct support for the developmental
theory. The most primitive feathers known—those
of Sinosauropteryx—are the simplest tubular structures
and are remarkably like the predicted stage 1
of the developmental model. Sinosauropteryx, Sinornithosaurus
and some other non-avian theropod
specimens show open tufted structures that lack a
rachis and are strikingly congruent with stage 2 of
the model. There are also pennaceous feathers that
obviously had differentiated barbules and coherent
planar vanes, as in stage 4 of the model."
Feduccia's frill argument was followed up in several other publications, in which researchers interpreted the filamentous impressions around Sinosauropteryx fossils as remains of collagen fibres rather than primitive feathers. Since the structures are clearly external to the body, these researchers have proposed that the fibres formed a frill on the back of the animal and underside of its tail, similar to some modern aquatic lizards.[20][21][22][23] The absence of feathers would refute the proposal that Sinosauropteryx is the most basal known theropod genus with feathers, and also raise questions about the current theory of feather origins itself. It calls into question the idea that the first feathers evolved not for flight but for insulation, and that they made their first appearance in relatively basal dinosaur lineages that later evolved into modern birds.[24]
Alleged primitive feathers or protofeathers in the theropod dinosaur Sinosauropteryxhave potentially profound implications concerning feather morphogenesis, evolution of flight, dinosaur physiology and perhaps even the origin of birds, yet their existence has never been adequately documented. We report on a new specimen of Sinosauropteryx which shows that the integumental structures proposed as protofeathers are the remains of structural fibres that provide toughness. The preservation in the proximal tail area reveals an architecture of closely associated bands of fibres parallel to the tail's long axis, which originate from the skin. In adjacent more exposed areas, the fibres are short, fragmented and disorganized. Fibres preserved dorsal to the neck and back and in the distal part of the tail are the remains of a stiffening system of a frill, peripheral to the body and extending from the head to the tip of the tail. These findings are confirmed in the holotype Sinosauropteryx and NIGP 127587. The fibres show a striking similarity to the structure and levels of organization of dermal collagen. The proposal that these fibres are protofeathers is dismissed.
In taxa more distantly related to birds, such as Sinosauropteryx(Figure 3 below), multiple tufts projecting a few millimeters from the skin have been discovered that resemble hypothesized early stages in avian feather development. These filamentous ‘feathers’ (or ‘protofeathers’; there is some disagreement concerning whether or not these integumentary structures were true feathers, e.g., Unwin 1998, Lingham-Soliar et al. 2007) were about 20 (5-40) mm long and appear to be rather homogenous over the body rather than originating in specific tracts. To some investigators, the filaments appear to be like down feathers and were probably used for insulation. They were hollow, and appeared to have a short shaft with barbs, but no barbules. In 2009, a fossil of another feathered dinosaur, Beipiaosaurus (a coelurosaurian theropod), with even simpler feathers was reported (Xu et al. 2009; Figures 4 and 5 below). These feathers consisted of single broad (about 2 mm wide) filament, were 10 to 15 centimeters long, and only present on the head, neck and tail. In taxa more closely related to birds, such as the oviraptorid Caudipteryx and dromaeosaurid Sinornithosaurus, elongate pinnate wing and tail feathers, structurally identical to the feathers of present-day birds and comprised of a central rachis, branching barbs, and barbules, have been found. In addition, fossils of a Dromaeosaurid (Microraptor) have revealed asymmetrically veined pennaceous feathers on both the forelimbs and hindlimbs (Clarke and Middleton 2006).
Spectacularly preserved non-avian dinosaurs with integumentary filaments/feathers have revolutionized dinosaur studies and fostered the suggestion that the dinosaur common ancestor possessed complex integumentary structures homologous to feathers. This hypothesis has major implications for interpreting dinosaur biology, but has not been tested rigorously. Using a comprehensive database of dinosaur skin traces, we apply maximum-likelihood methods to reconstruct the phylogenetic distribution of epidermal structures and interpret their evolutionary history. Most of these analyses find no compelling evidence for the appearance of protofeathers in the dinosaur common ancestor and scales are usually recovered as the plesiomorphic state,but results are sensitive to the outgroup condition in pterosaurs. Rare occurrences of ornithischian filamentous integument might represent independent acquisitions of novel epidermal structures that are not homologous with theropod feathers.
All taxa were scored for the presence/absence of epidermal scales, unbranched filaments (protofeathers)/quills and more complex branched filaments (including feathers).
The
“more complex
branched filaments” category contains taxa with compound filaments that are
not feathers and compound
filaments that are
feathers.
From the Supplementary
information:
The taxa with
compound branched filaments that are not feathers are Kulindadromeus, Dilong and
Ornithomimus.
The taxa with
compound branched filaments that are feathers begin at Pennaraptora.
This means that the
only intermediates between taxa with unbranched
filaments (protofeathers)/quills on the one hand and full pennaceous feathers in
Pennaraptora on the other are Kulindadromeus,
Dilong and Ornithomimus.
Most of our analyses provide no support for the appearance of feathers in the majority of non-avian dinosaurs, and although many meat-eating dinosaurs were feathered, the ancestor of all dinosaurs was probably scaly.
We considered attempting to describe the feather morphotypes in Kulindadromeus using the nomenclature of Prum et al. (52, 53) or of Xu et al. (21, 22). However, except for our monofilaments (which correspond well to Type 1 in Xu et al.), we could not assign with confidence the other two feather morphotypes in Kulindadromeus to categories described by Prum et al. or Xu et al. Further, fundamental discrepancies between these two previously published nomenclature systems remain to be resolved. Thus we felt that until new fossil material and a synthesis of existing nomenclature systems are available, interpretations of direct homologies between complex feather-types in Kulindadromeus and in Prum et al. or Xu et al. would be premature.
So I find it quite strange and disheartening that Godefroit et al.—despite being fairly objective in their supplementary material—go completely gung-ho in calling these structures feathers.
Old World porcupines (Hystricidae) have quills embedded in clusters, whereas in New World porcupines (Erethizontidae), single quills are interspersed with bristles, underfur, and hair.
The discovery in the late 1990s in China
of fossils from thousands of bona fide dinosaurs covered in
feathers provided the most definitive visual evidence for the
dinosaur–bird link [15–17], convincing most of the remaining
skeptics (Figure 2A–C). It is now widely accepted, even by ornithologists,
that birds evolved from dinosaurs [18], with the two
groups linked by hundreds of shared features of the skeleton,
soft tissues, growth, reproduction, and behavior [2,3,19–22].
Most amazingly, it is now known that many non-bird dinosaurs
were feathered and would have looked much more like birds
than lizards or crocodiles (Figure 3).
In my large-scale analysis of theropods (in preparation), Scansoriopterygidae are placed sligtly more basal than Avialae: they’re basal paravians, sister-group of Eumaniraptora (Avialae+Deinonychosauria). This different position explains more the “incisivosaur-like” skull and the absence of some pelvic features widespread among basal avialans, dromaeosaurids and troodontids (in particular the scapular, ischial and pubic features). In my blog, I suggested an alternative and very heterodox interpretation of Scansoriopterygids: given the absence of evidence for remiges in Epidexipteryx, is it possible that the “feather impressions” seen in the forelimb of the “Scansoriopteryx heilmanni specimen” are not feathers, but a different tegument: it is interesting to note that in remige-bearing maniraptorans, the remiges are inserted on the second finger, whereas in Scasoriopteryx these impression are close to the hyper-elongated third finger. This very long lateral finger is similar to the pterosaurian fourth digit. So, it is possible that the “feather impressions” of Scansoriopteryx were remnant of a patagium. In my opinion, the presence of this structure may explain the elongation of the lateral digit in these small theropods better than the Aye-Aye hypothesis.
Recent discoveries in Asia have greatly increased our understanding of the evolution of dinosaurs’ integumentary structures, revealing a previously unexpected diversity of “protofeathers” and feathers. However, all theropod dinosaurs with preserved feathers reported so far are coelurosaurs. Evidence for filaments or feathers in noncoelurosaurian theropods is circumstantial and debated. Here we report an exceptionally preserved skeleton of a juvenile megalosauroid, Sciurumimus albersdoerferi n. gen., n. sp., from the Late Jurassic of Germany, which preserves a filamentous plumage at the tail base and on parts of the body. These structures are identical to the type 1 feathers that have been reported in some ornithischians, the basal tyrannosaur Dilong, the basal therizinosauroid Beipiaosaurus, and, probably, in the basal coelurosaur Sinosauropteryx. Sciurumimus albersdoerferi represents the phylogenetically most basal theropod that preserves direct evidence for feathers and helps close the gap between feathers reported in coelurosaurian theropods and filaments in ornithischian dinosaurs, further supporting the homology of these structures. The specimen of Sciurumimus is the most complete megalosauroid yet discovered and helps clarify significant anatomical details of this important basal theropod clade, such as the complete absence of the fourth digit of the manus. The dentition of this probably early-posthatchling individual is markedly similar to that of basal coelurosaurian theropods, indicating that coelurosaur occurrences based on isolated teeth should be used with caution.
The best soft tissue preservation is found on the tail, which preserves large patches of skin, especially on the ventral but also on the dorsal side, and very fine, long, hair-like filaments that correspond to type 1 feathers (2) dorsally in the anterior midsection (Fig. 3 Cand D).
A
series of what appear to be hollow, tubular bristle-like structures,
approximately 16 centimetres (6.3 in) long, were also preserved, arranged in a
row down the [Psittacosaurus] dorsal(upper) surface of
the tail.
To put this in context,
they can be related to the Xu and Guo Morphotype 1:
Morphotype
1 is
known from the heterodontosauridTianyulong and the
ceratopsian Psittacosaurus (Mayr et
al., 2002; Zheng et al., 2009). Its main characteristic is being
monofilament
and relatively great length and rigidity. A variant
of this morphotype is seen in Beipiaosaurus, which differs from those of
Tianyulong and Psittacosaurus in its relatively great width ( Xu et al.,
2009b).Five major morphogenesis events are inferred to have
occurred sequentially in feather evolution before the origin of the Aves and
they are: 1) appearance of filamentous and tubular morphology, 2) formation of follicle
and barb ridges, 3) appearance of rachis, 4) appearance of
planar form, and 5) formation of pennaceous
barbules.
Note that it is only at
Morphotype 2 that the
follicle appears.
Morphotype 1 is a
bristle. For example, the bristles in Tianyulong
and Psittacosaurus.
We can see that shown
for Tianyulong
and Psittacosaurus in
Figure
5 of the Xu et al
study:
Here we report a new ornithischian dinosaur,
Kulindadromeus zabaikalicus, with diverse epidermal appendages, including
grouped filaments that we interpret as avianlike feathers.
But this is an overstatement since the Supplementary Information contains this: “We considered attempting to describe the feather morphotypes in Kulindadromeus using the nomenclature of Prum et al. (52, 53) or of Xu et al. (21, 22). However, except for our monofilaments (which correspond well to Type 1 in Xu et al.), we could not assign with confidence the other two feather morphotypes in Kulindadromeus to categories described by Prum et al. or Xu et al."
As we saw earlier,
Morphotype 1 is simply a bristle. So all that can be said with assurance, is
that Kulindadromeus had
bristles.
Filamentous integumentary structures are preserved in
all three [Yutyrannus] specimens. Those preserved in ZCDM V5000 are evidently associated with
the posterior caudal vertebrae. As preserved, they areparallel
to each otherand
form an angle of about 30u with the long axis of the tail. The filaments are at
least 15 cm long. They
are too densely packed for it to be possible to determine whether they are
elongate broad filamentous feathers (EBFFs) like those seen in the
therizinosauroid Beipiaosaurus, slender monofilaments, or compound filamentous
structures.Those
of ZCDM V5001 are near the pelvis and pes. They are filamentous structures, but
morphological details are not preserved.
All that can be said
with assurance, is that Yutyrannus had
bristles..
“The
fossil of a small, predatory dinosaur discovered in Germany has
experts rethinking how feathers developed among the dinosaurs that likely gave
rise to birds.The
authors say the new species underminesthe
notion that a covering of simple, hairlike feathers was characteristic of such
early theropods as was previously believed. Given
its position in the dinosaur family tree, Juravenator "should bear filamentous
feathers," Xing Xu said in an interview. But
Chiappe says the new fossil didn't seem to bear any physical evidence of
feathers, missing or not. "You could expect to see follicle [in the
skin], small pits that contain feather buds. We don't see them
in Juravenator,"
Chiappe said.”
A cladistic analysis indicates that the new [Juravenator] taxon is closer to maniraptorans than to tyrannosauroids, grouping it with taxa often considered to be compsognathids. Large portions of integument are preserved along its tail. The absence of feathers or feather-like structures in a fossil phylogenetically nested within feathered theropods5, 6 indicates that the evolution of these integumentary structures might be more complex than previously thought.
Portions of the [Juravenator] epidermis preserved mainly along the tail provide the only glimpse of the morphology of the skin of basal coelurosaurs, and structures newly revealed under UV light hint at the possibility of filamentous integumentary structures – akin to those interpreted as proto-feathers in other basal coelurosaurs – also covering the body of this dinosaur.
In 2014, Christian Foth and others argued that the evidence was insufficient to conclude that the forelimb feathers of Ornithomimus were necessarily pennaceous, citing the fact that the monofilamentous wing feathers in cassowaries would likely leave similar traces.[6]
Our reanalysis of two distinct phylogenetic datasets focusing on basal paravian taxa supports the reinterpretation of Balaur as an avialan more crownward than Archaeopteryx but outside of Pygostylia, and as a flightless taxon within a paraphyletic assemblage of long-tailed birds.
Based on phylogenetic analyses and critique of the various unusual features of this theropod, we argue that Balaur is likely not a dromaeosaurid, but a secondarily flightless bird. If you’re at all aware of the discussion that’s surrounded the possible evolution of flightlessness in non-bird paravians (Paul 1988, 2002), the significance of this won’t be lost on you....There’s more to say about our reinterpretation of Balaur. If a dromaeosaurid-like maniraptoran has turned out to be a flightless bird, does this have implications for any of the other flightless paravians of the Jurassic and Cretaceous?
Previously proposed hypothesis that flightless derived members of the maniraptoriform clade Deinonychosauria (Dinosauria: Theropoda) evolved from volant ancestors is evaluated by reviewing relevant publications subsequent to that of the hypothesis. Functional morphology and computer and physical modeling indicate that basal Dromaeosauridae microraptorine Microraptor and unenlagiine Rahonavis were volant, the former being capable of gliding and powered flight utilizing long pennaceous feathers on fore and hind limbs and scansorial locomotion, supporting the hypothesis that the more derived flightless dromaeosaurids evolved from volant ancestors.
I think dromaeosauridae's taxonomy will need a complete revision, Because the group is obviously too diversified and with a temporal and spacial range too wide to be fit in just a single family. There were smaller and bigger ones (some Giants), climbers, runners, gliders (flyers too?), Carnivores and omnivores (and herbivores?), From Last Last Jurassic to Cretaceous, in asiamerica, Europe, South America, Madagascar. I guess That All current "subfamilies" will be raised to families, Particularly Gondwanian Unenlagiinae, Whose evolution requires some degree of independent development and endemic. Their Paleogeography sounds a bit similar to multituberculate mammals, That Also had a massive presence in Asiamerican Cretaceoys, endemic offshoots in Europe and poorly known Gondwanian branches.
Yu, et al., did do some work with the suppression of genetic and molecular signaling pathways to see the effects on feather development. When they suppressed sonic hedgehog, they found that the resulting feather had barb rami partially joined by membranes.
To further test the role of SHH in feather branching, we suppressed SHH using cyclopamine27 or RCAS–antisense SHH in the plucked and regenerating feather model. The two independent reagents gave similar results. The regenerated feathers showed regions where barbs fused with a web-like membranebetween; therefore forming continuous feather vanes (Fig. 4b, c). Cross sections showed regions with barb ridges that failed to separate because the marginal plate cells failed to disappear (Fig. 4d, e). Suppressing SHH produced a similar phenotype as over-expressing BMP4 (compare Fig. 3g and and4e).4eand4e).4e).
Indeed, when neontological and palaeontological data illustrate almost all the series proposed by theories (Prum & Brush 2002; Xu 2006), transition remains particularly unclear before the stage III defined by Prum (1999), which is considered by Xu (2006)as probably the most critical stage of feather evolution in birds or non-avian dinosaurs.
The variation of space between adjacent actinofibrils in Jeholopterus, also reported in Rhamphorhynchus (Padian & Rayner 1993), suggests that those fibres were connected by some elastic tissue that enabled them to spread apart or join whenever necessary, making the actinopatagium more flexible (perhaps somewhat elastic
The hairs [pycnofibres] are shown to be complex multi-strand structures instead of single strands or actual hairs. The complex nature of these filaments most closely resembles natal down feathers, but apparently without having barbules. As such, they may represent the earliest known form of feathers.









v3.jpg)









