See also this link
http://pterosaurnet.blogspot.ca/2010/05/acetabulum.html
PELVIC BONES
http://icb.oxfordjournals.org/cgi/content/full/40/1/87#SEC3
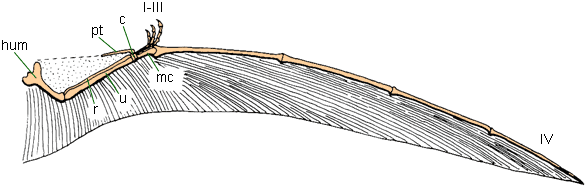
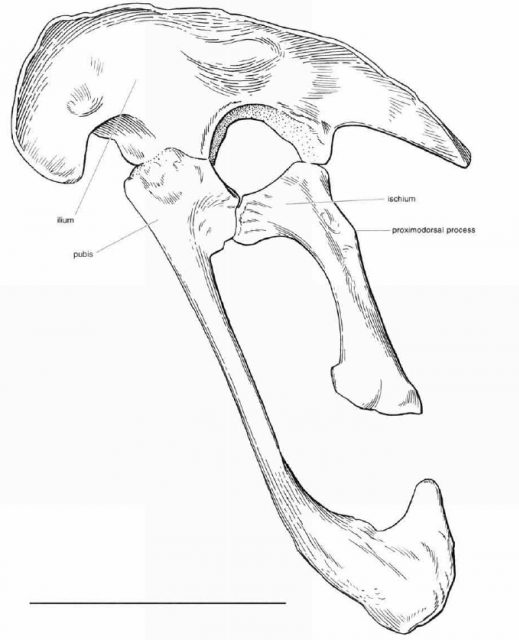
http://pterosaur.net/fossils_coloborhynchus_pelvis.php"Several aspects of the pelvic girdle suggest that pterosaurs were specialized for pelvic aspiration during flight. As in birds, the three pelvic bones of pterosaurs were solidly fused into a single unit (Fig. 9), and an increased number of dorsal vertebrae were incorporated into the sacrum (3 to 5 in Rhamphorhynchusand as many as 10 in Pteranodon) (Wellnhofer, 1978http://www.livescience.com/3399-north-american-dinosaur-smaller-housecat.html, 1987
"The claws were thought to come from juveniles — they were just so small. But when we studied the pelvis, we found the hip bones were fused, which would only have happened once the animal was fully grown," Longrich said. "Until now, the smallest carnivorous dinosaurs we have seen in North America have been about the size of a wolf. Judging by the amount of material that was collected, we believe animals the size of Hesperonychus [a dromaeosaurid] must have been quite common on the landscape."

The pelvis of the orinithocheiroid Coloborhynchus.http://pterosaur.net/fossils/coloborhynchus_pelvis_2.jpg
http://pterosaurnet.blogspot.ca/2010/05/pubic-bones.html
http://pterosaurnet.blogspot.ca/2012/12/no-connection-between-dinosaurs-and.html
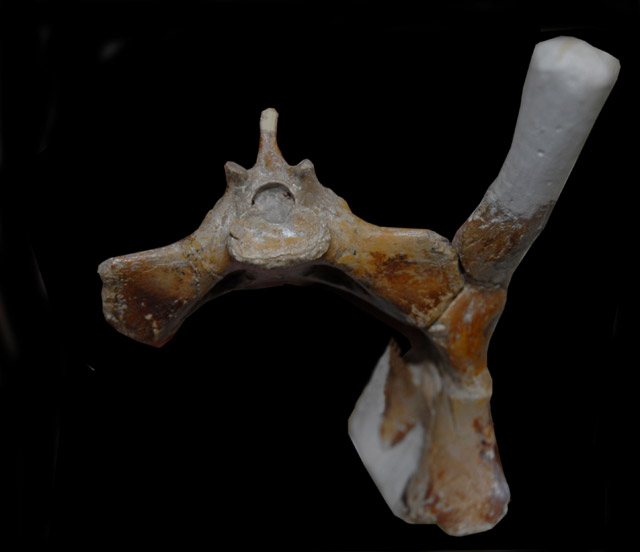
ACETABULUM/FEMUR ARTICULATION
SUMMARY:http://www.aou.org/auk/content/130/1/0001-0013.pdf
A partially closed acetabulum is seen in basal archosaurs and is characteristic of the scansoriopterids and Jurassic feathered forms such as Anchiornis initially described as near Aves by Xu et al (2009)
GENERAL:
See page 10
The hip joint of pterosaurs is more mobile and profoundly different from that of theropods but is reminiscent of that of mammals, especially of humans, allowing a wide range of adduction and abduction in the vertical plane.
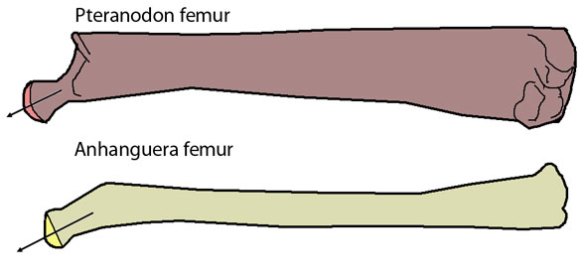
The femoral component [of the Anhanguera pterosaur femur] is a well defined spherical head which is distinctly separated from the shaft by a narrow non-articular neck at an obtuse angle of 160 degrees. The head forms a ball and socket joint with the close-fitting, shallow and imperforate acetabulum.
In theropods [dinosaurs], the femoral component is cylindrical without any distinctive head and neck. It projects medially at a right angle [90 degrees] from the shaft and fits into a perforated acetabulum of up to 1.5 times its diameter. As a result, the hip joint is stable and fully congruent during parasagittal motion, permitting a wide range of flexion and extension but very little abduction and adduction.
http://pterosaurheresies.wordpress.com/2011/10/14/pterosaur-femur-time/
Two pterodactylid pterosaurs:
http://www.lusofossils.com/verte.html
Dinosaur:

But notice the Anchiornis femur (d):
http://www.ivpp.ac.cn/qt/papers/201403/P020140314379333425828.pdf
http://scienceblogs.com/tetrapodzoology/wp-content/blogs.dir/471/files/2012/04/i-3f9364c23349a5c25c429724b628ee11-Anchiornis_details_Jan_2009.jpg
Anchiornis:

Bambiraptor:
http://kuscholarworks.ku.edu/dspace/...ries.013op.pdf
Hesperonychus:
http://www.pnas.org/content/106/13/5002.full

MODERN BIRD
Birds are the only living vertebrate whose hind limb includes three long bones in sequence. The innermost of the two long bones are similar to those found in most vertebrates. At the hip, the femur, is held more or less parallel to the ground and is bound to the hipbones by the massive thigh muscles. In effect, the femur is an addition to the hipbones and its rotation contributes little to the length of the bird’s stride.

During walking and running in birds, hindlimb movement is generated primarily at the knee and ankle joints; in humans, movement occurs at the knee, ankle and hip joints. The bird's thigh does not move substantially from its nearly horizontal position where it provides rigid lateral support to the thin walled air-sacs of the respiratory system. (Credit: Image courtesy of Oregon State University).
Note the pterosaur prepubis:
http://pterosaurheresies.wordpress.com/2011/11/06/whats-with-that-deep-prepubis/
The “dark wing” specimen of Rhamphorhynchus muensteri JME SOS 4785 (Tischlinger and Frey 2002) has one overlooked oddity worth mentioning. It had an incredibly deep prepubis (Figure 1.)
http://archosaurmusings.wordpress.com/2009/12/24/back-to-that-pterosaur-sacrum-pelvis
So much so normal, but pterosaurs also have a fourth pelvic bone in the form of the pre-pubis.http://www.bioone.org/doi/abs/10.1642/0004-8038%282007%29124%5B789%3ATAOBFA%5D2.0.CO%3B2?journalCode=tauk
The Auk 124(3):789–805, 2007
THE ANTITROCHANTER OF BIRDS: FORM AND FUNCTION IN BALANCE
Fritz Hertel1,3 and Kenneth E. Campbell, Jr.2
Abstract.—The antitrochanter is a uniquely avian osteological feature of the
pelvis that is located lateral to the postero-dorsal rim of the acetabulum. This feature
makes the avian hip joint unique among all vertebrates, living and fossil, in
that a significant portion of the femoral–pelvic articulation is located outside of the
acetabulum. This additional acetabular articulation occurs between the neck of the
femur and the antitrochanter, and operates as a hinge joint or ginglymus. It is complementary
to the articulation of the head of the femur with the acetabulum, which
is a pivot joint or trochoides. The size, location, and spatial orientation of the antitrochanter
were determined for 77 species of birds representing a variety of hindlimb
functions (e.g., highly cursorial, vertical clinging, foot-propelled diving) and spanning
a wide range of body sizes (swifts to rheas). The area of the antitrochanter is
a good predictor of body mass in birds; its position and orientation are reasonably
consistent within hindlimb morphofunctional groups, but not among all birds. The
antitrochanter serves as a brace to prevent abduction of the hindlimb and to absorb
stresses that would otherwise be placed on the head of the femur during bipedal
locomotion. The drum-in-trough-like form of the antitrochanter–femur articulation tends to assist in the transfer of long-axis rotational movements of the femur to the pelvis. The avian antitrochanter is a derived feature of birds that evolved as an aid in maintaining balance during bipedal terrestrial locomotion.
http://en.wikipedia.org/wiki/Pterosaur
Also:Pterosaurs' hip sockets are oriented facing slightly upwards, and the head of the femur (thigh bone) is only moderately inward facing, suggesting that pterosaurs had a semi-erect stance. It would have been possible to lift the thigh into a horizontal position during flight as gliding lizards do.
https://www.app.pan.pl/archive/published/app59/app20111109.pdf
Evolution of the pterosaur pelvis
https://en.wikipedia.org/wiki/Sacrum
The sacrum (/ˈsækrəm/ or /ˈseɪkrəm/; plural: sacra or sacrums;[1] Latin os sacrum[2]) in human anatomy is a large, triangular boneat the base of the spine, that forms by the fusing of sacral vertebrae S1–S5, between 18 and 30 years of age.[3]
In birds the sacral vertebrae are fused with the lumbar and some caudal and thoracic vertebrae to form a single structure called the synsacrum
https://en.wikipedia.org/wiki/Synsacrum
The synsacrum is a skeletal structure of birds and dinosaurs, in which the sacrum is extended by incorporation of additional fused or partially fused caudal or lumbar vertebrae.
http://www.prehistoriclife.xyz/history/mesozoic-birds.html
Archaeopteryx, as we have seen, had many features that are far from the condition found in living birds, including teeth, an unfused hand, a bony tail, no synsacrum, and gastralia.Subsequent events included the formation of a pygostyle as well as the development of the synsacrum and other features for a rigid trunk, all of which contribute to the efficient flight that characterizes modern birds.
Closer related yet to Aves were the toothed Ichthyornithiformes (see Figure 11.2). Unlike hesperornithiforms, ichthyornithiforms were excellent flyers (Figure 11.7). Ichthyornis, from the Late Cretaceous of North America, had a massive keeled sternum and an extremely large deltoid crest that was probably an adaptation for powerful flight musculature. In other respects, it shared many of the adaptations of modern birds including a shortened, fused trunk, a carpometacarpus, a pygostyle, a completely fused tarsometatarsus, and a synsacrum formed of 10 or more fused vertebrae. Found exclusively in marine deposits, ichthyornithiforms must have been rather like Mesozoic sea gulls - but with teeth.
http://www.reinhold-necker.de/seite14.html
The sacral enlargements of non-avian dinosaurs do not share much structural similarities with the lumbosacral specializations of modern birds. Furthermore these species were quadrupedal and had a long tail, i.e. there was no need for a sense organ of equilibrium. It seems that early birds with a pygostyle and a well- developped synsacrum show structural specializations which resemble that of modern birds. However, there is a need for more details and more examples of well- preserved fossils.
During evolution the reduction of the tail is accompanied by a melting of lumbosacral vertebrae named „synsacrum“.
http://bio.sunyorange.edu/updated2/pl%20new/55%20first%20birds.htm
Rahona ostromi lived in the late Cretaceous but its pelvic bones resemble those of the first birds. (Rahona means “menacing cloud.) It possessed a number of theropod characteristics (such as a pubic boot and the articulations between its vertebrae) but had the distinctive sickle claw and hyperextensible 2nd toe found in dinosaurs such as Velociraptor. It had primitive theropod characteristics such as a long tail, a saurischian vertebral articulation (also seen in Patagonykus), a pubic boot (as in Archaeopteryx and the enantiornithine birds) and some pelvic features (similar to Archaeopteryx and Unenlagia). It possessed a synsacrum which is an advanced feature (Forster, 1998; Gibbons, 1998).https://evodevojournal.biomedcentral.com/articles/10.1186/2041-9139-5-25
The oldest known bird, Archaeopteryx, dated to 150 million years ago, defines the clade Aves [5, 6, 7] or Avialae [8]. Its fully formed flight feathers, elongated wings, and evidence of capable powered flight, all ally Archaeopteryx with birds [9, 10]. Yet, the presence of teeth, clawed and unfused fingers, and an elongated, bony tail are characteristics shared with non-avian theropod dinosaurs. Paravians, including Archaeopteryx, are characterized by long tails [11, 12], some fusion of synsacral vertebrae, and varying flight capability (Figure 1). Most deinonychosaurians had between 20 and 30 caudal vertebrae. Oviraptorosaurs, probably the immediate outgroup to Paraves, had relatively shorter tails. These shorter tails were due not just to a modest decrease in the number of caudal vertebrae relative to other non-avian theropods, but more generally to a reduction in individual lengths of the more distal caudals [13]. Interestingly, several oviraptorosaurs have been documented to have the distal caudal vertebrae co-ossified into a pygostyle-like structure that braced a fan-like arrangement of retrices [13, 14, 15, 16]. Another more prominent independent reduction of tail length occurred in Epidexipteryx, a Mid- or Late-Jurassic maniraptoran dinosaur [17]. Its tail had only 16 caudal vertebrae with the distal ten tightly articulated to form a stiffened rod supporting four unique, ribbon-like, tail feathers.
https://static-content.springer.com/image/art%3A10.1186%2F2041-9139-5-25/MediaObjects/13227_2014_Article_146_Fig3_HTML.jpg

http://www.ivpp.ac.cn/qt/papers/201403/P020140314389417822583.pdf
The holotype specimen of Xiaotingia zhengi has completely closed neurocentral sutures on all exposed vertebrae and has a completely fused synsacrum, indicative of a late ontogenetic stage (probably adult).
Five sacral vertebrae form a short synsacrum (less than 60% as long as the ilium), as in other archaeopterygids and basal deinonychosaurs.
http://www.app.pan.pl/article/item/app20120093.html
W. Scott Persons, IV, Philip J. Currie, and Mark A. Norell
Oviraptorosaur caudal osteology is unique among theropods and is characterized by posteriorly persistent and exceptionally wide transverse processes, anteroposteriorly short centra, and a high degree of flexibility across the pre-pygostyle vertebral series. Three-dimensional digital muscle reconstructions reveal that, while oviraptorosaur tails were reduced in length relative to the tails of other theropods, they were muscularly robust. Despite overall caudal length reduction, the relative size of the M. caudofemoralis in most oviraptorosaurs was comparable with those of other non-avian theropods. The discovery of a second Nomingia specimen with a pygostyle confirms that the fused terminal vertebrae of the type specimen were not an abnormality. New evidence shows that pygostyles were also present in the oviraptorosaurs Citipati and Conchoraptor. Based on the observed osteological morphology and inferred muscle morphology, along with the recognition that many members of the group probably sported broad tail-feather fans, it is postulated that oviraptorosaur tails were uniquely adapted to serve as dynamic intraspecific display structures. Similarities, including a reduced vertebral series and a terminal pygostyle, between the tails of oviraptorosaurs and the tails of theropods widely accepted as basal members of the Avialae, appear to be convergences.
http://brantworks.com/resources/Allosaurus/ANTS_kit/ANTS-Allosaurus-Frank-Galef-detail-spine-BrantWorks.jpg

Feduccia
The (Archaeopteryx) synsacrum is also smaller and less fused, and the pubis is not fully reversed.
http://www.pterosaur.co.uk/PDB2012/P/index.htm
Pterosaurs all have a diapsid skull showing two openings behind the orbit. They have air sacs and pneumatic bones, a sclerotic ring around the eye and a distinctly fused synsacrum and pelvis.
The (Archaeopteryx) synsacrum consists of 5 fused vertebrae.