It is significant that all the core nodes in the hypothesized dinosaur to bird phylogeny are poorly supported.
For example:
Maniraptoriformes is poorly supported
Maniraptora is poorly supported.
Oviraptorosauria + Paraves (Pennaraptora) is poorly supported.
Paraves is poorly supported.
http://ib.berkeley.edu/courses/ib200a/labs/ib200a_lab10_bootstrap_jackknife_bremer.pdf
As a rule of thumb, a Bremer score of 3 is good and a score of 5 is “highly supported.”
Bootstrapping calculates a support value for each node based on the fraction of samples that support that node. The highest support value is 100, while values below 70 are usually considered weak. Values below 50 aren’t shown; in fact, branches below 50 are collapsed and shown as a polytomy.http://www.life.umd.edu/labs/delwiche/MSyst/lec/bootstrap.html
Low bootstrap values (below 50%) are essentially meaningless
Brusatte et al
http://www.cell.com/current-biology/fulltext/S0960-9822(14)01047-1 (2014)
https://www.cell.com/cms/10.1016/j.cub.2014.08.034/attachment/3d79921a-ae54-4bd9-b128-fa7140c7691f/mmc1.pdf (Supplemental Information)
across the Dinosaur-Bird Transition
Stephen L. Brusatte, Graeme T. Lloyd, Steve C. Wang, and Mark A. Norell
This clade—Maniraptoriformes—is only poorly supported (Bremer support of 1 and jackknife percentage of less than 50%), and relationships at its base are unresolved. There is a basal polytomy consisting of four clades: Ornitholestes, Compsognathidae, Ornithomimosauria, and Maniraptora (i.e., the clade of all taxa more closely related to birds than to Ornithomimus: [S52]).
Maniraptora—the clade defined as all taxa closer to birds than to Ornithomimus—is comprised in the present study of Alvarezsauroidea, Therizinosauroidea, Oviraptorosauria, and Paraves. This clade is supported by a Bremer value of 2 but a jackknife percentage of less than 50%.
The clade consisting of Oviraptorosauria and Paraves is supported by a Bremer value of 1 and a jackknife percentage of less than 50%.
Paraves—consisting of dromaeosaurids, troodontids, and avialans—is also poorly supported, as it also has a Bremer value of 1 and a jackknife of less than 50%.
Both Bremer supports and jackknife percentages (absolute values) were calculated in TNT using standard scripts. The jackknife was run using the default parameter of 36% character removal probability and 1000 replicates.
Note that there are enough characters in the study to make conclusions about all of the other major coelurosaurian subgroups so it is not a problem of not having enough data:
See Figure S1 for the large polytomy.
See below:
Numbers next to nodes denote Bremer support value/jackknife percentage. Those nodes without any numbers are characterized by Bremer values of 1 and a jackknife percentage of less than 60%
All of the major coelurosaurian subgroups that have long been considered monophyletic are also found to be monophyletic here. These include Tyrannosauroidea, Compsognathidae, Ornithomimosauria, Alvarezsauroidea, Therizinosauroidea, Oviraptorosauria, Dromaeosauridae, Troodontidae, and AvialaeAnd
Perhaps surprisingly, Ornithomimosauria is not found to be monophyletic and there is a large polytomy at the base of the clade that includes all coelurosaurs more derived (closer to avialans) than tyrannosauroids. This lack of resolution is due to the uncertain phylogenetic position of a small handful of taxa, including the fragmentary basal coelurosaur Kinnareemimus (a purported ornithomimosaur:Note that the core nodes are poorly supported even after the removal of those 5 taxa!
[S49]), the aberrant coelurosaur Epidendrosaurus (which is known only from two juvenile individuals: [S50]), the paravians Pyroraptor and Hesperonychus, and the avialan Limenavis. Pyroraptor and Limenavis were also found to be unstable in the analysis of Turner et al. (2012) [S9], whereas Epidendrosaurus was excluded from the primary version of that analysis
Ornithomimosaurs vs. tyrannosauroids: 29 axes, Mahalanobis distance=783720, p=0.00049975
The reduced strict consensus topology is shown in Figures S1-S2 (and Dryad Figs. S2-S3) and is used here as the preferred phylogeny and basis for character optimization and discussion of coelurosaurian phylogeny and evolution. This topology is considerably more resolved than the strict consensus, and recovers a monophyletic Ornithomimosauria and better resolution among basal coelurosaurs and paravians.http://datadryad.org/resource/doi:10.5061/dryad.84t75
See Figure S1 for the large polytomy.
See below:
Numbers next to nodes denote Bremer support value/jackknife percentage. Those nodes without any numbers are characterized by Bremer values of 1 and a jackknife percentage of less than 60%


Xu et al
https://www.researchgate.net/publication/51527707_An_Archaeopteryx-like_theropod_from_China_and_the_origin_of_Avialae (2011)
https://media.nature.com/original/nature-assets/nature/journal/v475/n7357/extref/nature10288-s1.pdf (Supplementary Information)
An Archaeopteryx-like theropod from China and the origin of Avialae
Xing Xu1,2, Hailu You3 , Kai Du4 & Fenglu Han2
It should be noted that our phylogenetic hypothesis is only weakly supported by the available data. Bremer support and bootstrap values for the recovered coelurosaurian subclades are, in general, low, and a bootstrap value less than 50% and a Bremer support value of 2 are obtained for a monophyletic Deinonychosauria including the Archaeopterygidae (see Supplementary Information). This low support is partly caused by various homoplasies, many of which are functionally significant, that are widely distributed across coelurosaurian phylogeny29.
We ran Bremer support and bootstrap analyses on the data matrix, using TNT with all default settings except that 1000 replications were used. Bremer support values for the recovered clades are indicated on Figure S8, and only clades with bootstrap values greater than 50% are shown in Figure S9. It is notable that only a few clades meet this criterion in the present analysis.Figure S9:

We evaluated the relative length and robustness of the forelimbs of major theropod groups by comparing the length and diameter of the humerus to the corresponding measurements for the femur in selected theropod taxa that represent the major theropod clades (Table S1). The plotted graph (Figure S4) indicates that the humerus is proportionally longer and more robust in basal avialans, archaeopterygids, and basal dromaeosaurids than in non-paravian theropods, troodontids, and derived dromaeosaurids. The last two groups are interpreted as having secondarily shortened forelimbs.
Figure S4. The relative length and diameter of the humerus in several theropod taxa. We use the ratios of humeral length to femoral length, and humeral diameter to femoral diameter, as indicators of forelimb length and robustness. Relative to the femur, the humerus is significantly longer and thicker in basal paravians than in non-paravian theropods, derived dromaeosaurids and troodontids (the relatively short and slender forelimbs in the last two groups are secondarily evolved according to the current phylogenetic analysis).
http://evolution.berkeley.edu/evolibrary/article/phylogenetics_03
Lack of knowledge
Usually, a polytomy means that we don't have enough data to figure out how those lineages are related. By not resolving that node, the scientists who produced the phylogeny are telling you not to draw any conclusions — and also to stay tuned: often gathering more data can resolve a polytomy.
Rapid speciation
Sometimes a polytomy means that multiple speciation events happened at the same time. In this case, all the daughter lineages are equally closely related to one another. The researchers who have reconstructed the tree you are examining should tell you if they feel that the evidence indicates that this is the case.
http://peter.unmack.net/molecular/programs/tnt.bootstrapping.html
This provides minimal instructions for running a bootstrap analysis.
http://www.sciencedirect.com/science/article/pii/S0960982214011385
...as the authors and others have noted, low support values indicate that many branches near the origin of birds remain unstable 1, 2, 4 and 5.
- 1
- | | |
- 2
- | |
- 4
- | |
- 5
https://www.researchgate.net/publication/305748962_Binary_Particle_Swarm_Optimization_Versus_Hybrid_Genetic_Algorithm_for_Inferring_Well_Supported_Phylogenetic_Trees
If, for example, you recover the same node through 95 of 100 iterations of taking out one character and resampling your tree, then you have a good idea that the node is well supported (your bootstrap value in that case would be 0.95 or 95%).
If we get low support, that suggests that only a few characters support that node, as removing characters at random from your matrix leads to a different reconstruction of that node.
https://en.wikipedia.org/wiki/Monophyly
Monophyletic groups are typically characterized by shared derived characteristics (synapomorphies).
Turner et al
http://digitallibrary.amnh.org/handle/2246/6352 (2012)
A review of dromaeosaurid systematics and paravian phylogeny. (Bulletin of the American Museum of Natural History, no. 371)
Turner, Alan H. (Alan Hamilton); Makovicky, Peter J.; Norell, Mark.
See Figures 66, 67, 68 (GC) and 69, 70, 71 (Bremer)
The results from the entire dataset reflect a wide range of support for nodes across the entire tree (figs. 66–68). Unsurprisingly, coelurosaurian monophyly is extremely well supported (GC 95) with little contradictory evidence. The basal Tyrannosauroidea clade is also well supported as is the less inclusive Tyrannosauridae node (GC 79 and 73, respectively).
Most of the intervening nodes between Proceratosaurus bradleyi, Ornithomimosauria, and derived maniraptorans have extremely low support (GC values between 2 and 12). This is neither surprising nor very informative given that most of these nodes collapse in the strict consensus topology of the phylogenetic analysis due to the labile positions of Proceratosaurus bradleyi, Dilong paradoxus, and Coelurus fragilis.
Maniraptora is poorly supported in the analysis (GC 5).
The monophyly of Paraves is poorly supported (GC 3) in part because of the placement of Epidexipteryx at the base of the clade. Analyses excluding Epidexipteryx find less contradictory data for the clade (GC 54).On the other hand:
Within Avialae, basal nodes show extremely high support (GC values between 92 and 75) with little contradictory data present.
Using raw frequencies is not recommended because there are cases in which groups lacking support have a frequency of 0.5 (Goloboff et al., 2003). GC frequencies are preferable because they reflect the balance between the amount of evidence that corroborates a given clade with the amount that falsifies that group.
Here are the GC values for the 4 groups we are looking at:
See Figures 66 and 67.
Maniraptoriformes:
Maniraptora:
Oviraptorosauria and Paraves: 0?
Paraves: 3
Analysis of the Turner et al study:
The exceptionally low values for Maniraptoriformes (2) and for Maniraptora (5) show that there is a break at that point. Anything beyond that is not attached to Coelurosauria.
For reference on GC:
http://www.ctoz.nl/cgi/t/text/text-idx?c=ctz;sid=f00ad86e4a09afab87790276f0f94154;idno=m7903a02;view=text;rgn=div2;cc=ctz;node=m7903a02%3A3.4
The difference in frequencies GC was chosen because it is calculated as the difference between the frequency in which a given group is retrieved in the jackknife replicates and the most frequent contradictory group (Goloboff et al., 2003). Absolute frequencies (the usual method of counting frequencies in jackknife or bootstrap analysis) do not distinguish between a group with a frequency of 0.6 that is never contradicted, and a group with a frequency of 0.6 that is contradicted with a frequency of 0.4. GC frequencies distinguish these two cases, giving lower support values to the second type of groups (Goloboff et al., 2003).https://www.academia.edu/4605777/Behaviour_of_resampling_methods_under_different_weighting_schemes_measures_and_variable_resampling_strengths
Goloboff et al. (2003a) proposed that what actually measures the support is not the absolute frequency (F), but the difference in frequency between a group and its most frequent contradictory group (GC, for ‘‘group present ⁄ contradicted’’). GC values of -1, 0 and 1indicate maximum contradiction, indifference, and max-imum support, respectively. GC is useful to measure strength of contradiction and to obtain support values for groups with positive but low support, which are otherwise not reported by methods using absolute frequencies (real groups with frequencies below 50% that are not retained in the majority consensus tree).
Cau et al
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4476167/
The phylogenetic affinities of the bizarre Late Cretaceous Romanian theropod Balaur bondoc (Dinosauria, Maniraptora): dromaeosaurid or flightless bird? (2015)
Andrea Cau,
 1 Tom Brougham,#2 and Darren Naish#2
1 Tom Brougham,#2 and Darren Naish#2Note that Coelurosauria and the derived clades have exceptionally low values.
Figure 5: Updated dataset of Brusatte et al. (2014).
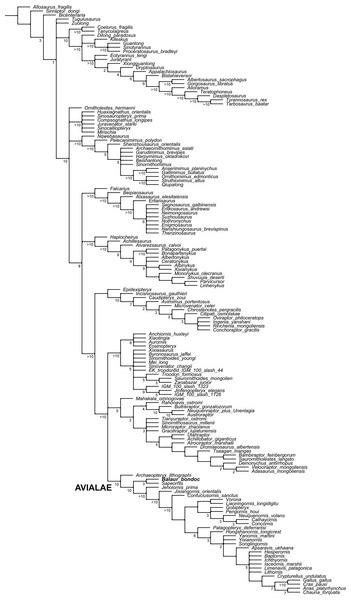
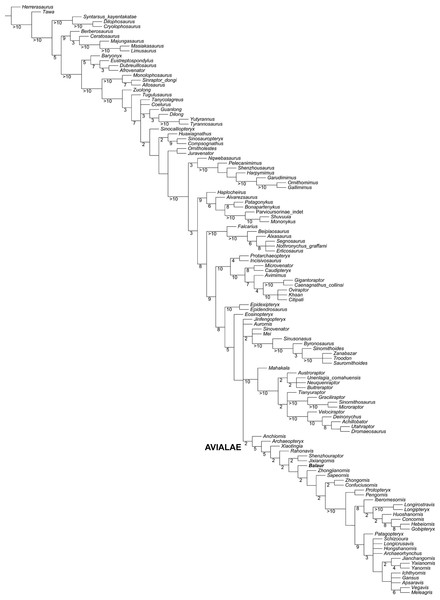
Figure 6: Updated dataset of Lee et al. (2014).
https://projecteuclid.org/euclid.ss/1063994980
http://projecteuclid.org/download/pdf_1/euclid.ss/1063994980
Pamela Soltis
Dinosaur to bird theorists claim that their purported dinosaur to bird lineage is the only solution available (given the data and the analytical method).
Abnormally rapid period of morphological evolution http://projecteuclid.org/download/pdf_1/euclid.ss/1063994980
Pamela Soltis
bootstrap values are low because of the small number of characters supporting each node
Dinosaur to bird theorists claim that their purported dinosaur to bird lineage is the only solution available (given the data and the analytical method).
But that is not correct.
See:
http://www.bio.fsu.edu/James/Ornithological%20Monographs%202009.pdf
AND of course the other possibility is that birds evolved from pterosaurs which is the idea being presented in this site.
=======================================================
See:
http://www.bio.fsu.edu/James/Ornithological%20Monographs%202009.pdf
AND of course the other possibility is that birds evolved from pterosaurs which is the idea being presented in this site.
=======================================================
Not only do the support indices indicate a problem, the dino to bird theory also requires a completely implausible rate of evolution. This is because there is a significant gap between paravians and non-paravians.
http://www.cell.com/current-biology/abstract/S0960-9822(14)01047-1
http://www.cell.com/current-biology/fulltext/S0960-9822(14)01047-1
Stephen Brusatte et al
Gradual Assembly of Avian Body Plan Culminated in Rapid Rates of Evolution across the Dinosaur-Bird Transition
recent studies converge in identifying the dinosaur-bird transition as an abnormally rapid period of morphological evolution.
Birds evolved significantly faster than other theropods, but they are indistinguishable from their closest relatives in morphospace.
The initial results of the branch (Dryad Fig. S4-13) and clade (Fig. S3; Dryad Fig. S14-23) tests strongly support significantly high rates in Avialae, and to a lesser degree Tyrannosauroidea.
Other clades show significantly low or non-significant rates, with the exception of two smaller clades: Graciliraptor + Microraptor + Shanag + Sinornithosaurus + Tianyuraptor (within Dromaeosauridae), and Anchiornis + Aurornis + Eosinopteryx + Xiaotingia (within Troodontidae), which frequently show high rates.
This provides robust evidence that birds (and their stem lineage) evolved faster than other theropods and that their origin was associated with an “early burst” of rapid morphological evolution. Previous studies have found significant changes in body size and limb morphology either progressively prior to the origin of birds or within more derived birds [3, 5, 6, 19, 20, 22], but our analysis of the overall phenotype puts the major rate shift at the origin of Avialae itself.
https://www.scientificamerican.com/article/how-dinosaurs-shrank-and-became-birds/
The ancestors of Paraves first started to shrink in size in the early Jurassic 200 million years ago, and fossil evidence show that this theropod line evolved new adaptations four times faster than other groups of dinosaurs,[8] and was shrinking 160 times faster than other dinosaur lineages were growing.[9]Xu, X.; Zhang, F. (2005). "A new maniraptoran dinosaur from China with long feathers on the metatarsus". Naturwissenschaften. 92 (4): 173–177. Bibcode:2005NW.....92..173X. doi:10.1007/s00114-004-0604-y. PMID 15685441.
A different set of tests in the Brusatte et al. [1] study compares rates between clades, revealing that birds as a clade exhibited a higher rate of skeletal evolution than other theropod clades.
http://www.ivpp.cas.cn/qt/papers/201403/P020140314389417822583.pdf
The discovery of Xiaotingia further demonstrates that many features
previously regarded as distinctively avialan actually characterize the
more inclusive Paraves. For example, proportionally long and robust
forelimbs are optimized in our analysis as a primitive character state
for the Paraves (see Supplementary Information). The significant
lengthening and thickening of the forelimbs indicates a dramatic shift
in forelimb function at the base of the Paraves, which might be related
to the appearance of a degree of aerodynamic capability.
http://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.915.8222&rep=rep1&type=pdf
http://science.sciencemag.org/content/sci/suppl/2014/07/30/345.6196.562.DC1/1252243.Lee.SM.revision1.pdf
Sustained miniaturization and anatomical innovation in the dinosaurian ancestors of birds
Michael S. Y. Lee,1,2* Andrea Cau,3,4 Darren Naish,5 Gareth J. Dyke5,6
Although there is no overall theropod-wide trend (fig. S7 and SM, part D), there is an exceptional trend within the single lineage that comprises much of the avian stem.
Our study quantifies rates of evolutionary innovation in dinosaurs using 1549 (data set 1) and 421 (data set 2) skeletal and other anatomical traits distributed across the entire body. A clear pattern emerges: Branches along the bird stem undergo substantially faster morphological evolution than those of the rest of the tree.
The theropod lineage directly ancestral to birds undergoes sustained miniaturization across 50 million years and at least 12 consecutive branches (internodes) and evolves skeletal adaptations four times faster than other dinosaurs.

=========================================================================
Ghost lineages
As if this is not bad enough this assumes millions of years of ghost lineages!
On the other hand:
https://en.wikipedia.org/wiki/Ornithomimosauria
Gregory S. Paul has proposed that Ornithomimosauria might be a group of primitive, flightless birds, more advanced than Deinonychosauria and Oviraptorosauria.[15]

Notice the ghost lineages:
https://www.researchgate.net/publication/269715801_An_integrative_approach_to_understanding_bird_origins

https://academic.oup.com/sysbio/article-abstract/42/2/182/1730933/An-Empirical-Test-of-Bootstrapping-as-a-Method-for
Bootstrapping is a common method for assessing confidence in phylogenetic analyses. Although bootstrapping was first applied in phylogenetics to assess the repeatability of a given result, bootstrap results are commonly interpreted as a measure of the probability that a phylogenetic estimate represents the true phylogeny.Brusatte bootstrap from TNT:
