ACETABULUM
Pterosaurs had a completely closed (cup-like) acetabulum that allowed them to abduct (splay, sprawl) their legs.
Dinosaurs had a completely open acetabulum that did not allow them to abduct (splay, sprawl) their legs.
DINOSAURS
http://books.google.ca/books?id=idta...lum%2C&f=false
Quote:
| In theropods, the femoral component is cylindrical without any distinctive head and neck. It projects medially at a right angle from the shaft and fits into a perforated [completely open] acetabulum of up to 1.5 times its diameter. As a result, the hip joint is stable and fully congruent during parasagittal motion, permitting a wide range of flexion and extension but very little abduction and adduction |
Quote:
| One important dinosaurian synapomorphy is the perforate [completely open] acetabulum, simply a "hip bone" (actually three connected bones, together called the pelvis) with a hole in the center where the head of the femur ("thigh bone") sits. This construction of the hip joint makes an erect stance (hindlimbs located directly beneath the body) necessary — like most mammals, but unlike other reptiles which have a less erect and more sprawling posture. Dinosaurs are unique among all tetrapods in having this perforate [completely open] acetabulum. |
BASAL PARAVES
http://ncsce.org/pdfs/home/aukfeduc2013.pdf
Quote:
| A number of intriguing four-winged feathered Jurassic
forms—such as the tiny scansoriopterids Epidendrosaurus
(= Scansoriopteryx) and Epidexipteryx, the latter without
preserved wing remiges, and anchiornithids (Anchiornis
and Xiaotingia)—exhibit numerous non-theropod skeletal features. ) They are provisionally best interpreted as early birds
at a pre-theropod stage, with partially closed hip joint or acetabulum,
and without a dinosaurian supra-acetabular shelf,
characters associated with a fully theropodan parasagittal
gait, which diagnose the clade. Although there is no reasonable
morphological definition of “theropod,” one sine qua non
for dinosaur status in general is the presence of a completely
open acetabulum, associated with the suite of changes seen in
posture and gait, by which a more upright posture is attained,
with a parasagittal hindlimb positioning (front to back axis). A partially closed acetabulum is seen in basal archosaurs and is characteristic of the scansoriopterids and Jurassic feathered forms such as Anchiornis initially described as near Aves by Xu et al (2009) |
Quote:
| The [Hesperonychus] acetabulum is similar to those of other dromaeosaurids in that it lacks a prominent supracetabular crest (30, 36). However, anteriorly, the contribution of the ilium to the acetabulum is broad, and the anterior rim projects strongly laterally, as it does in Unenlagia(36). The medial opening of the acetabulum is partially closed, as it is in other Dromaeosauridae (36). The [Hesperonychus] acetabulum opens dorsolaterally rather than laterally, as is the case in Velociraptor (38), suggesting the ability to partially abduct the hindlimbs. This morphology is of interest in light of proposals that Microraptor gui abducted its feathered hindlimbs to function as airfoils (24). |
Quote:
| Velociraptor mongoliensis had a pelvis with a characteristic pubis that pointed downward and forward at an angle toward the ischium. The acetabulum of V. mongoliensis opened dorsolaterally, indicating that it could abduct and adduct its hind limbs. This morphological characteristic demonstrates that the ancestors of V. mongoliensis were probably capable of flight and therefore the flightlessness of Velociraptor was secondarily lost (Longrich and Currie. 2009). |
Quote:
| Microraptors have been reconstructed in two distinctive models, the four-winged gliding model with sprawled hindlimb wings, by which it was originally described in Nature (Xu et al. 2003), and a dinosaurian bipedal model, or biplane model, by which it is reconstructed with the hindlimbs held beneath the body, incapable of sprawling, in other words, like a tiny T. rex. The problem,of course, is that there is absolutely no reason the hindlimbs could not have been sprawled, as is the case in flying squirrels (Glaucomys spp.), flying lemurs (Dermoptera), etc., and even falling cats. Too, the sprawled model performs superiorly inwind-tunnel experiments (Alexander et al. 2010), most specimensare preserved with a sprawled posture, and the wingclaws are adapted for trunk climbing (Burnham et al. 2011). In addition, it would be difficult to imagine how selection could produce elongate, asymmetric hindlimb flight remiges by the most current paleontological reconstructions, in which the hindlimbs are held in flight beneath the body in obligate bipedal fashion, with elongate hindlimb wing feathers trailing behind, simply slicing through the air (Balter 2012) |
Quote:
| Fossils of the remarkable dromaeosaurid Microraptor gui and relatives clearly show well-developed flight feathers on the hind limbs as well as the front limbs. No modern vertebrate has hind limbs functioning as independent, fully developed wings; so, lacking a living example, little agreement exists on the functional morphology or likely flight configuration of the hindwing. Using a detailed reconstruction based on the actual skeleton of one individual, cast in the round, we developed light-weight, three-dimensional physical models and performed glide tests with anatomically reasonable hindwing configurations. Models were tested with hindwings abducted and extended laterally, as well as with a previously described biplane configuration. Although the hip joint requiresthe hindwing to have at least 20° of negative dihedral (anhedral),all configurations were quite stable gliders. Glide angles rangedfrom 3° to 21° with a mean estimated equilibrium angle of 13.7°,giving a lift to drag ratio of 4.1:1 and a lift coefficient of 0.64. The abducted hindwing model’s equilibrium glide speed corresponds to a glide speed in the living animal of 10.6m·s−1. Although the biplane model glided almost as well as the other models, it was structurally deficient and required an unlikely weight distribution (very heavy head) for stable gliding. Our model with laterally abducted hindwings represents a biologically and aerodynamically reasonable configuration for this four-winged gliding animal. M. gui’s feathered hindwings, although effective for gliding, would have seriously hampered terrestrial locomotion. |
Quote:
| Scansoriopteryx also lacks a fully perforated acetabulum, the hole in the hip socket which is a key characteristic of Dinosauria and has traditionally been used to define the group. |
Quote:
| Scansoriopteryx is clearly more primitive than Archaeopteryx in many respects such as its saurischian-style pelvis which has remarkably short pubes; elongate and robust ischia; and comparatively small pubic peduncles. These primitive features further suggest that the nearly closed acetabulum is not a reversal, but a true plesiomorphic condition. |
Quote:
| This results in a somewhat sprawling position for the [Archaeopteryx] femur that is corrected at the knee joint, resulting in a functionally vertical leg. The pelvis has an incompletely open acetabulum, and there is no characteristic dinosaurian supra-acetabular shelf. The femoral head turns forwards rather than extending perpendicular to the shaft. |
http://books.google.ca/books?id=Sihl...osture&f=false
There is a reference to the splayed posture of Archaeopteryx here (page 399):
http://books.google.ca/books?id=8QRK...0angle&f=false
http://link.springer.com/article/10.1007%2Fs10336-014-1098-9#page-1
head of [Scansoriopteryx] femur lacks a distinctive neck and is instead more proximally oriented as in reptiles with sprawling limbs
http://books.google.ca/books?id=SihlpQTlVdAC&pg=PA154&lpg=PA154&dq=Certainly+the+fact+that+scansoriopterids&source=bl&ots=jTl6Uvn68h&sig=WRNGO1pzEQdzfw0F8HUmvrTHaVE&hl=en&sa=X&ei=ONVsVJ-GMI-nyASPzYCAAQ&ved=0CCEQ6AEwAA#v=onepage&q=Certainly%20the%20fact%20that%20scansoriopterids&f=false
Certainly the fact that scansoriopterids could spread the hind limbs outward in a splayed posture, more than in typical birds, indicates that a true upright stance was achieved only later and independently from true dinosaurs.Also see the info about splayed hindlimbs here and here.
the [Scansoriopteryx] acetabulum is not as fully perforated as in any known theropod
Jurassic archosaur is a non-dinosaurian bird
Stephen A. Czerkas, Alan Feduccia
Unlike theropod dinosaurs, invariably exhibiting aPTEROSAUR
completely perforated and open acetabulum, Scansoriopteryx
has a partially closed acetabulum, and no sign of a
supra-acetabular shelf or an antitrochanter. Along with the
mostly enclosed acetabulum indicated by the surface texture
of the bone within the hip socket, the proximally
oriented head of the femur is functionally concordant with
a closed or partially closed acetabulum and with sprawling
hindlimbs. There is additional phylogenetic evidence that
the largely closed acetabulum was not directly inherited
from dinosaurian ancestors with fully open acetabulae and
subsequentially modified as a secondary reversal. The
similar condition seen in Anchiornis (Hu et al. 2009) and
Microraptor (personsal observations; Xu et al. 2000; Gong
et al. 2012) with the partially open acetabulum in
Scansoriopteryx creates a sequential phylogenetic pattern
consistent with being inherited from non-dinosaurian
archosaurs which had not yet achieved a fully upright
stance as in dinosaurs (Fig. 2).
A fully perforated acetabulum is a sine qua non for
dinosaurian status associated with major changes in posture
and gait, by which a more upright posture and parasagittal
stance is attained.
The pterosaur acetabulum was completely closed because during development the ilium, ischium and pubis extended to come completely together at the bottom of the cup-shaped acetabulum.
In the transition to basal paraves, the ilium, ischium and pubis no longer came completely together. (ie. it was partially open). This may well be an example of neoteny.
http://pterosaur.net/terrestrial_locomotion.php
Now, this is not to say that basal pterosaurs were locomotory inept from the moment they landed. They may, however, have spent more time running around trees and cliffs than over floodplains and tidal flats. Basal pterosaurs typically have deepened, highly recurved manual and pedal claws with comparatively large flexor tubercles compared to the relatively slender claws of pterodactyloids. These claws are extremely thin despite their depth and would make excellent crampons to provide purchase when climbing, especially when combined with the antungual sesamoids and elongate penultimate phalanges that characterise the hands and feet of many basal forms. Furthermore, the orientation of the femoral head in basal pterosaurs means that the femur is projected forward, upward and laterally from the acetabulum, thereby causing the sprawling gait for the hindlimbs that acted in concert with the relatively short metacarpals to bring the bodies of these pterosaurs close to any surface they happened to be climbing over. These are all excellent adaptations to climbing (Fig. 5), and we should expect early Mesozoic environments to be covered with pterosaurs hanging from cliff faces, tree trunks and branches.Lunate Surface
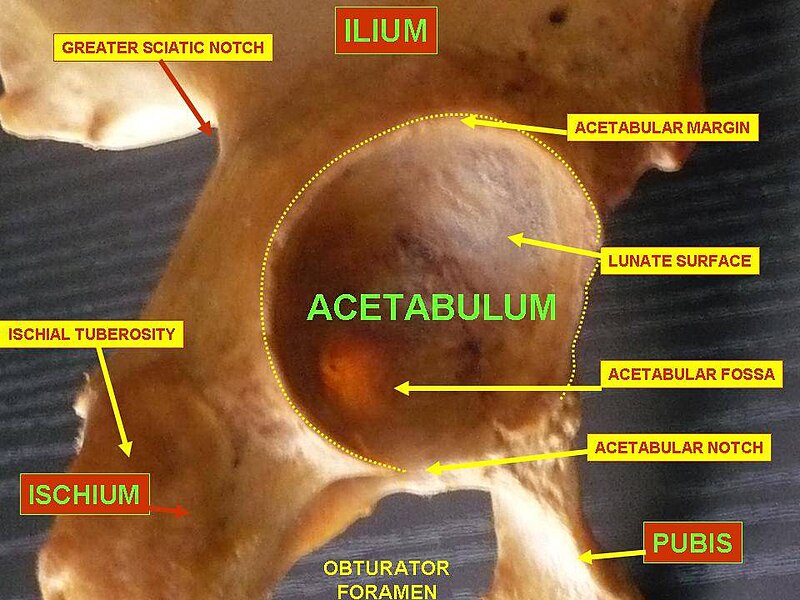
https://en.wikipedia.org/wiki/Acetabulum
The rest of the acetabulum is formed by a curved, crescent-moon shaped surface, the lunate surface, where the joint is made with the head of the femur.
Its counterpart in the pectoral girdle is the glenoid fossa.[3]
Related links:
http://pterosaurnet.blogspot.ca/2010/05/acetabulum.html
http://pterosaurnet.blogspot.ca/2013/01/pelvic-bones-summary.html
What is the function of the pelvis? – The pelvis connects the hind limbs to the trunk of the body (through the acetabulum). In Dinosaurs, a robust ridge [supra-acetabular shelf, crest, ridge] present above the acetabulum on the ilium helps transmit the weight of the animal to the hind limb.
The [dinosaur] parasagittal limb posture was achieved by making the process (bump) on the head of the femur (thigh bone) that articulates with the hip socket (or acetabulum) protrude at a right angle from the rest of the femur, which as a result will point directly downward.However, this process is cylindrical, not ball-like as for most vertebrates. This means that the femur cannot roll around along the joint, but is restricted to rotating only. As a result, the leg can only be swung forward and backward, and not to the sides as, for example, we humans can. In addition, the ankle joint of the dinosaurs was relatively simple: in humans, there are seven ankle bones (or tarsals), and they form a synovial joint, a highly flexible joint that allows movement in several planes; dinosaurs, however, only have two ankle bones, and the joint (referred to as the mesotarsal joint) forms a linear hinge joint, which only enables movement in a single plane only, in this case back and forward. Consequently, the dinosaur hindlimb had a highly restricted range of movement: it could only move along the sagittal plane – the line parallel to the body. Thus, dinosaurs would not have been very agile regarding sidewise movement; they were specialised for running forward.https://sites.google.com/site/dinolore/_/rsrc/1318797976741/learn/dinosaur-groups/dinosauria/Sprawling_and_erect_hip_joints_-_horiz.png

https://en.wikipedia.org/wiki/Acetabulum
The well-fitting surfaces of the femoral head and acetabulum, which face each other, are lined with a layer of slippery tissue called articular cartilage, which is lubricated by a thin film of synovial fluid. Friction inside a normal hip is less than one-tenth that of ice gliding on ice.[4][5]
No comments:
Post a Comment