Here is a first draft list of changes that would have taken place from
Rhamphorhynchidae (pterosaur) to Scansoriopterygidae (basal paraves):
Large 4th finger reduced
Patagium greatly reduced
Pycnofibres developed into wing flight feathers
Uropatagium replaced with flight feathers
Distal syncarpal becomes semilunate carpal and fuses to metacarpals
The pteroid bone and lateral carpal are lost
Acetabulum is not completely closed
Second toe becomes hyper-extended
5th toe is lost?
Clavicles no longer incorporated into the sternum
Humerus saddle-shaped head becomes bulbous
External mandibular fenestra appeared
Posterior teeth reduced
First toe is turned backwards?
If anyone would care to suggest a change or addition, please do. Just provide reference link(s) and copy and paste the material that you think supports your suggestion.
NEW
The discovery of Yi qi would change the transition steps*.
The major changes would be:
The large 4th finger is first reduced to just the metacarpal (in Yi) [or last phalange] and then that is also lost (in other scansoriopteryids).
The remaining digits would be digits 1, 2 and 3. (The first digit would not be lost, the 4th digit would be lost).
* if the interpretation of the Yi characteristics by Xu et al is correct.
Saturday, September 27, 2014
Monday, September 22, 2014
Rhamphorhynchidae to Scansoriopterygidae
I suggest that Rhamphorhynchidae (basal pterosaur group) is the ancestor of Scansoriopterygidae (basal paraves group).
Scansoriopterygidae are basal paraves with flight feathers on the arms and legs.
Scansoriopterygidae (or a group very much like it) is the ancestor of the later Paraves such as Microraptor, Archaeopteryx etc.
Rhamphorhynchidae pycnofibres are homologous (ancestral) to Scansoriopterygidae feathers.
The Rhamphorhynchidae long bony tail is homologous to the Scansoriopterygidae long bony tail.
The Rhamphorhynchidae caudal rods are homologous to the Scansoriopterygidae caudal rods.
The Scansoriopterygidae elongate outermost digit is transitional between Rhamphorhynchidae and later Paraves.
The Scansoriopterygidae wing feathers replaced the function of the wing membrane of their pterosaur ancestor. The membrane close to the arm (patagium) remained.
The Scansoriopterygidae hindwing feathers replaced the function of the uropatagium of their pterosaur ancestor.
The Scansoriopterygidae propatagium IS the propatagium from their pterosaur ancestor.
Scansoriopterygidae is one of the most basal (primitive) members of paraves.
It used the same muscles as pterosaurs for flying. It could splay its hind limbs like pterosaurs for flying.
The evidence strongly supports the transition from pterosaur to basal paraves, with Scansoriopterygidae being transitional.
http://en.wikipedia.org/wiki/Scansoriopterygidae

Scansoriopterygidae are basal paraves with flight feathers on the arms and legs.
Scansoriopterygidae (or a group very much like it) is the ancestor of the later Paraves such as Microraptor, Archaeopteryx etc.
Rhamphorhynchidae pycnofibres are homologous (ancestral) to Scansoriopterygidae feathers.
The Rhamphorhynchidae long bony tail is homologous to the Scansoriopterygidae long bony tail.
The Rhamphorhynchidae caudal rods are homologous to the Scansoriopterygidae caudal rods.
The Scansoriopterygidae elongate outermost digit is transitional between Rhamphorhynchidae and later Paraves.
The Scansoriopterygidae wing feathers replaced the function of the wing membrane of their pterosaur ancestor. The membrane close to the arm (patagium) remained.
The Scansoriopterygidae hindwing feathers replaced the function of the uropatagium of their pterosaur ancestor.
The Scansoriopterygidae propatagium IS the propatagium from their pterosaur ancestor.
Scansoriopterygidae is one of the most basal (primitive) members of paraves.
It used the same muscles as pterosaurs for flying. It could splay its hind limbs like pterosaurs for flying.
The evidence strongly supports the transition from pterosaur to basal paraves, with Scansoriopterygidae being transitional.
http://en.wikipedia.org/wiki/Scansoriopterygidae

Friday, September 19, 2014
Flight Strokes Compared
Basal Paraves used the same flight stroke as Pterosaurs and both had splayed hindlimbs for flying.
FLIGHT STROKE
PTEROSAURS
http://press.princeton.edu/witton/sampler-pterosaurs.pdf
http://biology.kenyon.edu/courses/biol241/bird%20flight%202003%20Chatterjee_Sankar.pdf
Pterosaurs and basal paraves flapped their wings in the same way.
___________________________________________________________
SPLAYED HINDLIMBS
Basal paraves had a femur/acetabulum articulation that was different than dinosaurs and that allowed them to abduct (splay) their legs.
See also these links
http://pterosaurnet.blogspot.ca/2010/05/acetabulum.html
http://pterosaurnet.blogspot.ca/2013/01/pelvic-bones-summary.html
PTEROSAUR
http://en.wikipedia.org/wiki/Pterosaur
Riddle of the Feathered Dragons
http://books.google.ca/books?id=SihlpQTlVdAC&pg=PA154&lpg=PA154&dq=Certainly+the+fact+that+scansoriopterids&source=bl&ots=jTl6Uvn68h&sig=WRNGO1pzEQdzfw0F8HUmvrTHaVE&hl=en&sa=X&ei=ONVsVJ-GMI-nyASPzYCAAQ&ved=0CCEQ6AEwAA#v=onepage&q=Certainly%20the%20fact%20that%20scansoriopterids&f=false
There is a reference to the splayed posture of Archaeopteryx here (page 399):
The Origin and Evolution of Birds
http://books.google.ca/books?id=8QRKV7eSqmIC&pg=PA399&lpg=PA399&dq=archaeopteryx+femur+angle&source=bl&ots=fqR1hR9GAi&sig=7grokjyLDiND0WyvJNaAJC8e0Ko&hl=en&sa=X&ei=p1UYVJW0H5OtyATf5YCYCA&ved=0CB0Q6AEwAA#v=onepage&q=archaeopteryx%20femur%20angle&f=false
http://www.bioone.org/doi/pdf/10.1525/auk.2013.130.1.1
Alexander et al
http://bioweb.uwlax.edu/bio203/s2014/gibbs_eliz/adaptation.htm
(Longrich and Currie. 2009)

Also interesting:
http://dml.cmnh.org/2005Dec/msg00079.html
That the leg winged dromaeosaurs have modified femoral heads that in some ways resemble those of some pterosaurs suggests they had evolved a different function, later lost in most secondarily flightless examples. Which is why so many specimens are splayed out spread eagle style. The hip sockets and leg musculature would not have to be fully functional when splayed out in flying dromaeosaurs because they were not using them for walking or running (many joints are not in full articulation in animals when they are not bearing full loads). (G. Paul)
Consider the following. We can see that they are assuming a dino to bird theory when concluding it could not splay its legs.
http://blogs.scientificamerican.com/tetrapod-zoology/2013/11/18/flight-of-the-microraptor/
Very good reference on pterosaur hip structure.
http://books.google.ca/books?id=idta6AVV-tIC&pg=PA10&lpg=PA10&dq=pterosaur+acetabulum+hip+socket&source=bl&ots=2GUU4Y1aSx&sig=f5JZsHtyLQHlEb3P4A9BIshxIkE&hl=en&sa=X&ei=DiMeVPrNCZG2yATXwICQCQ&ved=0CDcQ6AEwAw#v=onepage&q=pterosaur%20acetabulum%20hip%20socket&f=false
Related references:
http://www.sciencedaily.com/releases/2010/01/100125173238.htm
Pterosaur pelvis
http://icb.oxfordjournals.org/content/early/2011/09/21/icb.icr112.full
"Tent" model:
http://www.pnas.org/content/107/7/2733.full.pdf+html
John Ruben
http://www.pnas.org/content/106/13/5002.abstract?ijkey=b80c6cec04b5f05bcbf67870ea44df19a403ed62&keytype2=tf_ipsecsha
Hesperonychus
http://www.pnas.org/content/107/40/E155.full
Argument (Brougham and Brusatte)
Reply to Brougham and Brusatte
___________________________________________________________
The idea that flapping must have been first
The trees down theory begins with a gliding arboreal creature. This is closer to being correct except that the gliding phase took place much earlier in the evolution of the pterosaur. This is a key point.
http://blogs.scientificamerican.com/WSS/post.php?blog=43&post=9336
https://www.google.ca/url?sa=t&rct=j&q=&esrc=s&source=web&cd=1&cad=rja&uact=8&ved=0CB0QFjAA&url=http%3A%2F%2Fwww.springer.com%2Fcda%2Fcontent%2Fdocument%2Fcda_downloaddocument%2F9781461473961-c1.pdf%3FSGWID%3D0-0-45-1409215-p175092519&ei=6FMvVMmLGeKbjALf04CQDg&usg=AFQjCNHWM-i3q0hYliEZtZSLr-UJWlm40w&sig2=7sv3satbA1kcbbROg0H6dw&bvm=bv.76802529,d.cGE
A FANCIFUL RATIONALIZATION:
The passage below offers up a very weak rationalization for thinking that even if the trees down theory is correct the arboreal creatures could still be dinosaurs. As the writers admit, "It would require stages in the origin of flight not preserved in our current sample of extinct nonavian dinosaurs."
http://az.oxfordjournals.org/content/amzoo/40/4/486.full.pdf
AMER. ZOOL., 40:486–503 (2000)
Phylogenetic Context for the Origin of Feathers1
STUART S. SUMIDA2,* AND CHRISTOPHER A. BROCHU†
FLIGHT STROKE
PTEROSAURS
http://press.princeton.edu/witton/sampler-pterosaurs.pdf
Detailed reconstruction of the proximal armhttp://eurekamag.com/research/019/478/morphological-evolution-pectoral-girdle-pterosaurs-myology-role.php (2003)
musculature of pterosaurs shows that ... the arm was more likely
lifted by large muscles anchored on the scapula and
back, and lowered by those attached to the sternum
and coracoid (fig. 5.8; Bennett 2003a). Unlike [modern] birds,
where two vastly expanded muscles are mainly used
to power flight, it appears that pterosaurs used several
muscle groups to form their flapping strokes.
The musculature of the pectoral region of representative rhamphorhynchoid (Campylognathoides) and large pterodactyloid (Anhanguera) pterosaurs was reconstructed in order to examine the function of various muscles and the functional consequences of the evolution of the advanced pectoral girdle of large pterodactyloids. The reconstructions suggest that m. supracoracoideus was not an elevator of the wing, but instead depressed and flexed the humerus. m. latissimus dorsi, m, teres major, m. deltoides scapularis, and m. scapulohumeralis anterior were wing elevators.BASAL PARAVES
http://biology.kenyon.edu/courses/biol241/bird%20flight%202003%20Chatterjee_Sankar.pdf
Its lack of a supracoracoideus (SC)http://www.ncbi.nlm.nih.gov/pmc/articles/PMC1780066/
pulley, the primary elevator of the wing, would prevent
Archaeopteryx from executing humeral rotation on the
glenoid during the upstroke,
Anatomical evidence indicates that Microraptor was not capable of ground or running takeoff, because it lacked the supracoracoideus pulley to elevate the wings.http://jeb.biologists.org/content/200/23/2987.full.pdf
The lack of a morphologically derived SC in Late
Jurassic and Early Cretaceous birds precluded a high velocity
recovery stroke which undoubtedly limited
powered flight in these forms. Subsequent evolution of the
derived SC capable of imparting a large rotational force to
the humerus about its longitudinal axis was an important
step in the evolution of the wing upstroke and in the ability
to supinate (circumflex) the manus in early upstroke, a
movement fundamental to reducing air resistance during
the recovery stroke.
The highly derived morphology of the SC,https://www.google.ca/url?sa=t&rct=j&q=&esrc=s&source=web&cd=1&cad=rja&uact=8&ved=0CB4QFjAA&url=https%3A%2F%2Frepository.si.edu%2Fbitstream%2Fhandle%2F10088%2F6524%2FVZ_93_Archaeopteryx.pdf%3Fsequence%3D1&ei=uUF2VZyMFcqlyASSo4LYCQ&usg=AFQjCNFW5HcrkvSBGaZvOkczCMZAErCEow&sig2=0o1CnMU3Vz-DQ8_S-sw-Bw&bvm=bv.95039771,d.aWw
a characteristic of modern birds capable of powered flight, was
not present in Archaeopteryx (Ostrom, 1976a,b; Wellnhofer,
1988, 1993), nor is there firm evidence for its presence in
recently described Mesozoic species (Chiappe, 1995; Sanz et
al. 1996).
Pterosaurs and basal paraves flapped their wings in the same way.
___________________________________________________________
SPLAYED HINDLIMBS
Basal paraves had a femur/acetabulum articulation that was different than dinosaurs and that allowed them to abduct (splay) their legs.
See also these links
http://pterosaurnet.blogspot.ca/2010/05/acetabulum.html
http://pterosaurnet.blogspot.ca/2013/01/pelvic-bones-summary.html
PTEROSAUR
http://en.wikipedia.org/wiki/Pterosaur
"Pterosaur's hip sockets are oriented facing slightly upwards, and the head of the femur (thigh bone) is only moderately inward facing, suggesting that pterosaurs had a semi-erect stance. It would have been possible to lift the thigh into a horizontal position during flight as gliding lizards do."
BASAL PARAVES
http://en.wikipedia.org/wiki/Microraptor
There is a reference to the splayed posture of Scansoriopterids here (page 154):http://en.wikipedia.org/wiki/Microraptor
Some paleontologists have doubted the biplane hypothesis, and have proposed other configurations. A 2010 study by Alexander et al. described the construction of a lightweight three-dimensional physical model used to perform glide tests. Using several hind leg configurations for the model, they found that the biplane model, while not unreasonable, was structurally deficient and needed a heavy-headed weight distribution for stable gliding, which they deemed unlikely. The study indicated that a laterally abducted hindwing structure represented the most biologically and aerodynamically consistent configuration for Microraptor.[3] A further analysis by Brougham and Brusatte, however, concluded that Alexander's model reconstruction was not consistent with all of the available data on Microraptor and argued that the study was insufficient for determining a likely flight pattern for Microraptor. Brougham and Brusatte criticized the anatomy of the model used by Alexander and his team, noting that the hip anatomy was not consistent with other dromaeosaurs. In most dromaeosaurids, features of the hip bone prevent the legs from splaying horizontally; instead, they are locked in a vertical position below the body. Alexander's team used a specimen of Microraptor which was crushed flat to make their model, which Brougham and Brusatte argued did not reflect its actual anatomy.[15] Later in 2010, Alexander's team responded to these criticisms, noting that the related dromaeosaur Hesperonychus, which is known from complete hip bones preserved in three dimensions, also shows hip sockets directed partially upward, possibly allowing the legs to splay more than in other dromaeosaurs.[16]
Riddle of the Feathered Dragons
http://books.google.ca/books?id=SihlpQTlVdAC&pg=PA154&lpg=PA154&dq=Certainly+the+fact+that+scansoriopterids&source=bl&ots=jTl6Uvn68h&sig=WRNGO1pzEQdzfw0F8HUmvrTHaVE&hl=en&sa=X&ei=ONVsVJ-GMI-nyASPzYCAAQ&ved=0CCEQ6AEwAA#v=onepage&q=Certainly%20the%20fact%20that%20scansoriopterids&f=false
Certainly the fact that scansoriopterids could spread the hind limbs outward in a splayed posture, more than in typical birds, indicates that a true upright stance was achieved only later and independently from true dinosaurs.
There is a reference to the splayed posture of Archaeopteryx here (page 399):
The Origin and Evolution of Birds
http://books.google.ca/books?id=8QRKV7eSqmIC&pg=PA399&lpg=PA399&dq=archaeopteryx+femur+angle&source=bl&ots=fqR1hR9GAi&sig=7grokjyLDiND0WyvJNaAJC8e0Ko&hl=en&sa=X&ei=p1UYVJW0H5OtyATf5YCYCA&ved=0CB0Q6AEwAA#v=onepage&q=archaeopteryx%20femur%20angle&f=false
http://www.bioone.org/doi/pdf/10.1525/auk.2013.130.1.1
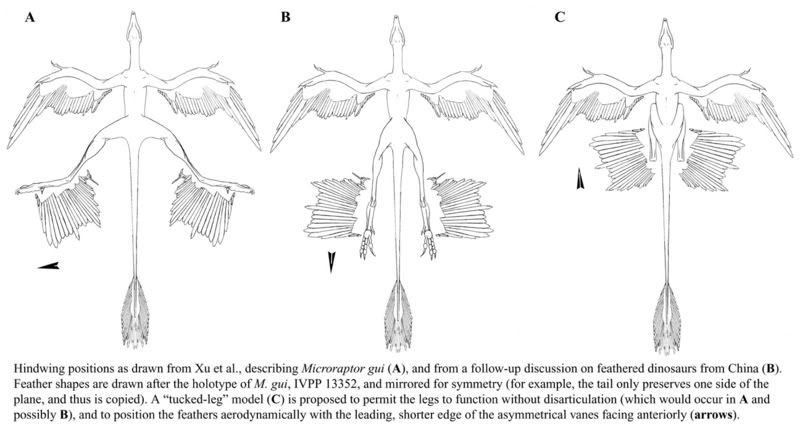
Microraptors have been reconstructed in two distinctive models, the four-winged gliding model with sprawled hindlimb wings, by which it was originally described in Nature (Xu et al. 2003), and a dinosaurian bipedal model, or biplane model, by which it is reconstructed with the hindlimbs held beneath the body, incapable of sprawling, in other words, like a tiny T. rex. The problem,of course, is that there is absolutely no reason the hindlimbs could not have been sprawled, as is the case in flying squirrels (Glaucomys spp.), flying lemurs (Dermoptera), etc., and even falling cats. Too, the sprawled model performs superiorly inwind-tunnel experiments (Alexander et al. 2010), most specimensare preserved with a sprawled posture, and the wingclaws are adapted for trunk climbing (Burnham et al. 2011). In addition, it would be difficult to imagine how selection could produce elongate, asymmetric hindlimb flight remiges by the most current paleontological reconstructions, in which the hindlimbs are held in flight beneath the body in obligate bipedal fashion, with elongate hindlimb wing feathers trailing behind, simply slicing through the air (Balter 2012)http://www.pnas.org/content/107/7/2972.full.pdf+html
Alexander et al
Fossils of the remarkable dromaeosaurid Microraptor gui and relatives clearly show well-developed flight feathers on the hind limbs as well as the front limbs. No modern vertebrate has hind limbs functioning as independent, fully developed wings; so, lacking a living example, little agreement exists on the functional morphology or likely flight configuration of the hindwing. Using a detailed reconstruction based on the actual skeleton of one individual, cast in the round, we developed light-weight, three-dimensional physical models and performed glide tests with anatomically reasonable hindwing configurations. Models were tested with hindwings abducted and extended laterally, as well as with a previously described biplane configuration. Although the hip joint requires the hindwing to have at least 20° of negative dihedral (anhedral),all configurations were quite stable gliders. Glide angles ranged from 3° to 21° with a mean estimated equilibrium angle of 13.7°,giving a lift to drag ratio of 4.1:1 and a lift coefficient of 0.64. The abducted hindwing model’s equilibrium glide speed corresponds to a glide speed in the living animal of 10.6m·s−1. Although the biplane model glided almost as well as the other models, it was structurally deficient and required an unlikely weight distribution (very heavy head) for stable gliding. Our model with laterally abducted hindwings represents a biologically and aerodynamically reasonable configuration for this four-winged gliding animal. M. gui’s feathered hindwings, although effective for gliding, would have seriously hampered terrestrial locomotion.
Primitively, early archosaurs are sprawling, with the legs set
laterally and elevated at around 75° (6), a preadapted posture for
gliding.
Modern birds normally have the thigh elevated and sprawled to the side in different degrees; for example, it is nearly perpendicular to the midline in loons and grebes (7).
This variation shows that the degree of splaying needed to use the
hindlegs in gliding is not unusual when compared with that in
modern birds. The absence of an antitrochanter and a supraacetabular
acetabular shelf (SAC) in the eumaniraptorans, including dromaeosaurids,
would make elevation and splaying of the legs even
easier (8). Air pressure could have provided most of the force
needed to elevate the leg into a gliding position similar to that in
gliding mammals. This simple positioning was originally assumed
for the four-winged Microraptor gui (5); but later, workers hoping
to recover an upright posture proposed arrangements of the
hindlimbs that would have required complicated systems of locks
and muscles to hold the leg in an only partially elevated position,
e.g., the “biplane” model (9). New anatomical information based
on the discovery of several hundred specimens similar to the
four-winged glider M. gui (and related taxa) has produced converging
lines of evidence demonstrating that the original
describers of M. gui (5) were correct in their interpretation of the
flight posture. We postulate, based on examination of this new
material, that M. gui was capable of abducting the hind limbs at
least 65–70° to achieve a gliding posture.
http://bioweb.uwlax.edu/bio203/s2014/gibbs_eliz/adaptation.htm
Velociraptor mongoliensis had a pelvis with a characteristic pubis that pointed downward and forward at an angle toward the ischium. The acetabulum of V. mongoliensis opened dorsolaterally, indicating that it could abduct and adduct its hind limbs. This morphological characteristic demonstrates that the ancestors of V. mongoliensis were probably capable of flight and therefore the flightlessness of Velociraptor was secondarily lost (Longrich and Currie. 2009).http://www.pnas.org/content/106/13/5002.full
(Longrich and Currie. 2009)
The acetabulum is similar to those of other dromaeosaurids in that it lacks a prominent supracetabular crest (30, 36). However, anteriorly, the contribution of the ilium to the acetabulum is broad, and the anterior rim projects strongly laterally, as it does in Unenlagia(36).
The medial opening of the acetabulum is partially closed, as it is in other Dromaeosauridae (36). The [Hesperonychus] acetabulum opens dorsolaterally rather than laterally, as is the case in Velociraptor (38), suggesting the ability to partially abduct the hindlimbs. This morphology is of interest in light of proposals that Microraptor gui abducted its feathered hindlimbs to function as airfoils (24).GENERAL
Possible shape:
http://pterosaurheresies.wordpress.com/2012/11/01/microraptor-leg-feathers-and-the-evolution-of-bird-flight/
Also interesting:
http://dml.cmnh.org/2005Dec/msg00079.html
That the leg winged dromaeosaurs have modified femoral heads that in some ways resemble those of some pterosaurs suggests they had evolved a different function, later lost in most secondarily flightless examples. Which is why so many specimens are splayed out spread eagle style. The hip sockets and leg musculature would not have to be fully functional when splayed out in flying dromaeosaurs because they were not using them for walking or running (many joints are not in full articulation in animals when they are not bearing full loads). (G. Paul)
Consider the following. We can see that they are assuming a dino to bird theory when concluding it could not splay its legs.
http://blogs.scientificamerican.com/tetrapod-zoology/2013/11/18/flight-of-the-microraptor/
One debate that surrounds the aerodynamic performance of Microraptor concerns hindlimb posture. In the very first study to discuss Microraptor’s possible flight abilities, it was depicted as being capable of a full-on sprawl, its hindlimbs projecting laterally in parallel with its arms (Xu et al. 2003). This sprawling pose was also promoted in another study (Alexander et al. 2010).
Given that the form of the theropod femur and hip socket generally prevents the hindlimb from being abducted this far from the sagittal plane (there are proximally placed trochanters on the femur, and supra-acetabular shelves and antritrochanters on the ilium that prevent this sort of posture), this is surely incorrect (Brougham & Brusatte 2010). http://www.nature.com/ncomms/2013/130918/ncomms3489/full/ncomms3489.html
Very good reference on pterosaur hip structure.
http://books.google.ca/books?id=idta6AVV-tIC&pg=PA10&lpg=PA10&dq=pterosaur+acetabulum+hip+socket&source=bl&ots=2GUU4Y1aSx&sig=f5JZsHtyLQHlEb3P4A9BIshxIkE&hl=en&sa=X&ei=DiMeVPrNCZG2yATXwICQCQ&ved=0CDcQ6AEwAw#v=onepage&q=pterosaur%20acetabulum%20hip%20socket&f=false
Related references:
http://www.sciencedaily.com/releases/2010/01/100125173238.htm
A joint team from the University of Kansas and Northeastern University in China says that it has settled the long-standing question of how bird flight began.http://www.app.pan.pl/archive/published/app59/app20111109.pdf
Pterosaur pelvis
http://icb.oxfordjournals.org/content/early/2011/09/21/icb.icr112.full
"Tent" model:
http://www.pnas.org/content/107/7/2733.full.pdf+html
John Ruben
http://www.pnas.org/content/106/13/5002.abstract?ijkey=b80c6cec04b5f05bcbf67870ea44df19a403ed62&keytype2=tf_ipsecsha
Hesperonychus
http://www.pnas.org/content/107/40/E155.full
Argument (Brougham and Brusatte)
The hypothesis that birds are theropod dinosaurs is supported by anatomical and molecular similarities, shared growth dynamics and physiology, and fossil theropods covered in feathers. A recent paper by Alexander et al. (1) and an associated commentary by Ruben (2) attempted to understand one of the greatest remaining riddles of avian evolution: the origin of flight itself.http://www.pnas.org/content/107/40/E156.full
Alexander et al. (1) examined the aerodynamic capabilities of Microraptor, a Cretaceous dromaeosaurid (avian sister taxon), by subjecting a reconstructed model to glide tests (1). They concluded that Microraptor was an arboreal glider that used all limbs as a single airfoil. We applaud the empirical approach of the study by Alexander et al.(1) and agree that Microraptor was capable of gliding. We disagree, however, with their anatomical reconstruction of Microraptor and, most importantly, with the assumption that any discovery about the habits of a single dromaeosaurid may solve the riddle of the origin of avian flight.
Alexander et al. (1) reconstructed Microraptor as a sprawling animal, with femora oriented at 140° to one another, a pelvic anatomy unlike that of other dromaeosaurids. The authors claimed that “new anatomical information,” gleaned from “examination of new material,” supported their reconstruction (1). However, this is simply asserted, with no description or illustration of this new material. Similarly, the authors argued that the lack of a supracetabular crest and an antitrochanter in Microraptor and other dromaeosaurids allows for a greater range of motion in the hind limb (1). This is incorrect: dromaeosaurids actually have enlarged antitrochanters, which limit femoral abduction (3), and although the supracetabular crest is reduced, it is still present (3). Finally, one author (J.B.) examined the cast pelvis used by Alexander et al. (1) and found that it, like all known Microraptor specimens, was crushed flat. With this in mind, it is important that the recent discovery of a 3D pelvis of a close relative, Hesperonychus (4), seems to allow only minor lateral splaying of the hind limbs.
We disagree with Ruben (2) on his presumption that different postural reconstructions of Microraptor “imply profoundly different scenarios for the origin of flight” (2). The implicit assumption is that this single species can be analyzed biomechanically, and whatever configuration glides best when launched from a catapult is the probable anatomy of the ancestor of birds. There is a clear fallacy in this reasoning: Microraptor itself cannot be an ancestor of birds, because it lived after birds had originated. It could only help understand avian flight if it retained the gliding abilities of that ancestor, which is not at all certain (5). There are nearly 40 known dromaeosaurid and troodontid dinosaurs—the closest relatives to birds. These animals exhibit a wide range in morphology, body size, integumentary covering, limb proportions, and inferred habitat. Fixation on a single derived dromaeosaurid species is not the path to understanding the origins of avian flight. We do commend Ruben (2), however, on acknowledging that an arboreal theropod dinosaur may have given rise to birds, which departs from his previous criticism of the dinosaur–bird hypothesis and is in line with the robustly supported theory that birds are living theropods.
Reply to Brougham and Brusatte
Brougham and Brusatte (1) agree with us that Microraptor was an arboreal glider but disagree with the posture of our model's hind limbs (1). They offer no suggestion for an alternative, other than the implied parasagittal posture of a typical dromaeosaur, which we showed was aerodynamically and mechanically unlikely (2). They cast doubt on the sprawled (abducted) hind-limb posture of our reconstruction—a key feature—by claiming that dromaeosaurid hips have structures (antitrochanters and supracetabular crests) that prevent abduction and that our specimen was too flattened to see such features. Whereas large dromaeosaurids may possess such structures, apparently, in small ones such as Microraptor, these structures are greatly reduced (3). The authors (1) misrepresent the small dromaeosaurid Hesperonychus as providing evidence against a sprawling posture, when the original description of Hesperonychus specifically mentions the lack of processes on the acetabulum preventing abduction and states that the “acetabulum opens dorsolaterally rather than laterally… suggesting the ability to partially abduct the hind limbs. This morphology is of interest in light of proposals that Microraptor gui abducted its feathered hind limbs to act as airfoils” (reference 4 in ref. 1). Their crushed flat claim is based on examining an incomplete cast (1). They did not examine the original fossil or its X-rays and X-ray computed tomography scans, and thus made a judgment using incomplete information. This Microraptor pelvis has been figured and described (4), and a complete 3D cast of the specimen is also available in our collections for examination. Furthermore, our pelvic morphology was checked against dozens of other specimens (as stated in ref. 2). We stand by our anatomical observations and are currently describing this specimen, along with other material, which will support the accuracy of our interpretation.
Brougham and Brusatte (1) devote a large portion of their letter to defending the dinosaurian origin of birds. We find this somewhat puzzling, because we did not address that issue in our paper. They choose to regard this feathered dromaeosaur as a derived member of the group, although most cladistic analyses show it as basal (5) or even as the primitive sister group of that clade (3). Moreover, much older taxa with large, pennaceous feathers on the lower hind limb (Pedopenna, Anchiornis) have been discovered from radiometrically dated Jurassic rocks in China (5). Anchiornis is cladistically a troodontid, suggesting that four-winged gliding is also primitive for deinonychosaurs. Brougham and Brusatte (1) suggest that Microraptor is of no relevance to understanding bird flight, because they doubt that it inherited its mode of gliding from an ancestor that it shared with birds. We think that such a shared ancestry is actually reasonable given the feathered hind limbs of Anchiornis, to which other authors have attributed an aerodynamic function (5). We never argued that Microraptor must be a direct ancestor of birds to be informative about the origin of avian flight, any more than Archaeopteryx must be ancestral to modern birds to be informative about avian origins. We think Microraptor displays a four-winged mode of gliding that it inherited from more primitive, arboreal ancestors, and we are confident that our model is anatomically reasonable.
___________________________________________________________
The trees down theory begins with a gliding arboreal creature. This is closer to being correct except that the gliding phase took place much earlier in the evolution of the pterosaur. This is a key point.
http://blogs.scientificamerican.com/WSS/post.php?blog=43&post=9336
Kevin Padian of the University of California at Berkeley, an expert on bird evolution, observed that the presentations focused on the effect of the hindlimb on a gliding animal instead of one that flapped its wings. Last year at the SVP meeting he presented evidence that gliders and flyers are completely unrelated to each other. He says that “there is not a shred of evidence that says gliding is involved in the evolution of flapping flight.” He questioned why the team's model would focus on gliding parameters when the forelimb shape was consistent with flapping, not gliding, and the hindlimb would have generated so much drag.http://www.economist.com/blogs/babbage/2011/11/evolution-flight
Rather, it was a challenge to the discipline's long-held belief that flapping wings, with their complicated nerve, muscle and bone structure, must, surely, have evolved from a simpler variety that allowed its owners to glide.
At first, Dr Padian and Dr Dial were expecting to find fossilised bats that were gliders, not flappers. To their surprise, they discovered nothing of the sort. They then expanded their search to all flying and gliding animals. What they found was, if anything, even more shocking: gliding and flapping fauna appear to have no direct common ancestor. This suggests that the prevailing [trees down] theory is wrong. It also, however, leaves open the question of where powered flight comes from.http://gwawinapterus.wordpress.com/2014/02/21/on-the-origins-of-flight-no-gliders/
An alternative option is what is reflected by an examination of gliding animal groups: gliding simply does not translate into flight, simply never leads to powered volancy, and that the flying vertebrates we know and love, as well as insects, have simply evolved under extremely anomalous sets of behaviour completly unrelated to gliding.
Based on birds and bats, as well as modern gliders, gliding appears to be effectively independent from flight, gliding animals seemingly unable to produce powered flight and animals that have evolved powered flight having developed it in unusual circumstances. Exceptions might exist, including possibly pterosaurs, but as it stands vertebrate flight has only been able to evolve from behaviours that promote forelimb movement, with aerodynamic speciation not leading at all to volancy.http://www.nature.com/news/ancient-bats-got-in-a-flap-over-food-1.9304
“It is a simple but amazing observation that there are no flying lineages of vertebrates with gliders as a sister group,” says evolutionary biologist Nancy Simmons at the American Museum of Natural History in New York.More:
https://www.google.ca/url?sa=t&rct=j&q=&esrc=s&source=web&cd=1&cad=rja&uact=8&ved=0CB0QFjAA&url=http%3A%2F%2Fwww.springer.com%2Fcda%2Fcontent%2Fdocument%2Fcda_downloaddocument%2F9781461473961-c1.pdf%3FSGWID%3D0-0-45-1409215-p175092519&ei=6FMvVMmLGeKbjALf04CQDg&usg=AFQjCNHWM-i3q0hYliEZtZSLr-UJWlm40w&sig2=7sv3satbA1kcbbROg0H6dw&bvm=bv.76802529,d.cGE
A FANCIFUL RATIONALIZATION:
The passage below offers up a very weak rationalization for thinking that even if the trees down theory is correct the arboreal creatures could still be dinosaurs. As the writers admit, "It would require stages in the origin of flight not preserved in our current sample of extinct nonavian dinosaurs."
http://az.oxfordjournals.org/content/amzoo/40/4/486.full.pdf
AMER. ZOOL., 40:486–503 (2000)
Phylogenetic Context for the Origin of Feathers1
STUART S. SUMIDA2,* AND CHRISTOPHER A. BROCHU†
The cursorial hypothesis, in its modern incarnation, is the simplest model for the origin of flight in the context of the preferred phylogenetic hypothesis. But is it a necessary corollary, as implied by some authors? We can infer the mechanism for the origin of flight from a phylogenetic tree, but this will always be a secondary inference. A strict reading of current cladograms does not necessarily reject a trees-down model of flight origins—the fossil record is incomplete, and one can always posit an unpreserved arboreal dinosaurian relative of birds. Additionally, rejection of a ground-up model for flight origins need not imply rejection of the dinosaurian hypothesis (Gauthier and Padian, 1985; Padian and Chiappe, 1998a), and some authors who derive birds from theropods explicitly prefer a trees-down model for flight origins (e.g., Chatterjee, 1999). The origin of a group, the origin of a structure, and the origin of a behavior or function are fundamentally different questions, and a cladogram primarily addresses the first. Some authors find legitimate reason to prefer a less-parsimonious scenario for a give cladogram on the basis of some sort of external evidence (e.g., Zaher and Rieppel, 1999).
One could argue that nonavian maniraptorans could climb, or that small arboreal theropods are yet to be discovered. Such a model would be much less parsimonious than the cursorial model, at least from the perspective of present phylogenetic understanding. It would require stages in the origin of flight not preserved in our current sample of extinct nonavian dinosaurs. The arboreal model is thus less parsimonious, but is not strictly falsified by the cladogram in Figure 1; an arboreal nonavian theropod may await discovery somewhere. The statement by Geist and Feduccia (2000) that cladistic analyses posit that “avian flight necessarily developed within a terrestrial context” (our emphasis) reflects a misunderstanding of how cladograms are constructed and what they can actually falsify.
Tuesday, September 16, 2014
Flight without a supracoracoideus
http://books.google.ca/books?id=8QRKV7eSqmIC&pg=PA136&lpg=PA136&dq=dorsal+elevator+feduccia&source=bl&ots=fqR1hQfECg&sig=NAYAvTNh7USVZlO-dFmaLcJsDPQ&hl=en&sa=X&ei=UUUYVIndD8-zyATm3oDgDg&ved=0CCQQ6AEwAQ#v=onepage&q=dorsal%20elevator%20feduccia&f=false
(Olson, Feduccia 1979)
Basal Paraves (feathered pterosaurs) were fliers. Just not able to take off from level ground.
The dorsal elevators, principally the deltoideus major, can effect the recovery stroke by themselves, as they did in Archaeopteryx. The German anatomist Maxheinz J. Sy proved this when he cut the tendons of the supracoracoideus in living crows and pigeons (1936). Sy found that pigeons were capable of normal, sustained flight; the only capacity they lost was the ability to take off from level ground. (Feduccia)https://repository.si.edu/bitstream/handle/10088/6524/VZ_93_Archaeopteryx.pdf?sequence=1&isAllowed=y
(Olson, Feduccia 1979)
Furthermore, the supracoracoideus muscle,
and hence an ossified sternum, is not necessary to effect the
recovery stroke of the wing. Thus the main evidence for
Archaeopteryx having been a terrestrial, cursorial predator is
invalidated. There is nothing in the structure of the pectoral girdle of Archaeopteryx that would preclude its having been a powered flier.
Basal Paraves (feathered pterosaurs) were fliers. Just not able to take off from level ground.
Friday, September 12, 2014
Four-winged flyers
An interesting video, which unfortunately is biased to the dino to bird theory.
Especially note from 48:00 to 49:00.Note that at 48:17 they point to exactly the spot where we find Scansoriopteryx.
https://www.youtube.com/watch?v=9-YNW_z_FUE
And particularly see 27:00 to 28:00.
At 27:37 notice the lineage that Larry Martin proposes that links archosaurs to basal paraves.
In what I am suggesting, that line is a branch of PTEROSAURS.
Also see here for the extra section that was not televised, of flight with splayed legs.
http://www.pbs.org/wgbh/nova/microraptor/extras.html
"The lift climbs steadily and more predictably than anything they've seen so far".
MICRORAPTOR
http://en.wikipedia.org/wiki/Microraptor#Wings_and_flight
Microraptor had four wings, one on each of its forelegs and hind legs. The long feathers on the legs of Microraptor were true flight feathers as seen in modern birds, with asymmetrical vanes on the arm, leg, and tail feathers. As in modern bird wings, Microraptor had both primary (anchored to the hand) and secondary (anchored to the arm) flight feathers. This standard wing pattern was mirrored on the hind legs, with flight feathers anchored to the upper foot bones as well as the upper and lower leg. It has been proposed that the animal glided and probably lived mainly in trees, because the hind wings anchored to the feet of Microraptor would have hindered their ability to run on the ground.[9] It had long pennaceous feathers on arms and hands (10–20 centimetres long or 3.9–7.9 in), with legs and feet 11–15 cm long (4.3–5.9 in). Toward the tail end, Microraptor was covered in shorter downy (plumulaceous) feathers, 2–6 cm long (0.79–2.36 in). Though not apparent in most fossils under natural light, due to obstruction from decayed soft tissue, the feather bases extended close to or in contact with the bones, as in modern birds, providing strong anchor points.[10]
When describing specimens originally referred to the distinct species Cryptovolans pauli, paleontologist Stephen Czerkas argued that Microraptor may have been able to fly better than Archaeopteryx, noting the fused sternum and asymmetrical feathers of Microraptor, as well as features of the shoulder girdle that indicate flying ability closer to modern birds than to Archaeopteryx. Czerkas cited the fact that this possibly volant animal is also very clearly a dromaeosaurid, to suggest that the Dromaeosauridae might actually be a basal bird group, and that later, larger, species such as Deinonychus were secondarily flightless. The work of Xu and colleagues also suggested that basal dromaeosaurs were probably small, arboreal, and could at least glide, though later discoveries of even more primitive dromaeosaurids with short forelimbs unsuitable for gliding have cast doubt on this view.[9][11]
Whether or not Microraptor could achieve powered flight or only passive gliding has been controversial. While most researchers have agreed that Microraptor had most of the anatomical characteristics expected in a flying animal, some studies have suggested that the shoulder joint was too primitive to have allowed flapping. The ancestral anatomy of theropod dinosaurs has the shoulder socket facing downward and slightly backward, making it impossible for the animals to raise their arms vertically, a prerequisite for the flapping flight stroke in birds. Some studies of maniraptoran anatomy have suggested that the shoulder socket did not shift into the bird-like position of a high, upward orientation close to the vertebral column until relatively advanced avialans like the enantiornithes appeared.[12] However, other scientists have argued that the shoulder girdle in some paravian theropods, including Microraptor, is curved in such a way that the shoulder joint could only have been positioned high on the back, allowing for a nearly vertical upstroke of the wing. This possibly advanced shoulder anatomy, combined with the presence of a propatagium linking the wrist to the shoulder (which fills the space in front of the flexed wing and may support the wing against drag in modern birds) and an alula or "bastard wing" may indicate that Microraptor was capable of true, powered flight.[13]
Chatterjee also used computer algorithms that test animal flight capacity to test whether or not Microraptor was capable of true, powered flight, in addition to passive gliding. The resulting data showed that Microraptor did have the requirements to sustain level powered flight, so it is theoretically possible that the animal flew on occasion in addition to gliding.[5]
Scansoriopterygidae
http://en.wikipedia.org/wiki/Scansoriopteryx
One distinctive feature of Scansoriopteryx is its elongated third finger, which is the longest on the hand, nearly twice as long as the second finger (in most theropod dinosaurs, the second finger is the longest). This is unlike the configuration seen in most other theropods, where the second finger is longest. The long wing feathers, or remiges, appear to attach to this long digit instead of the middle digit as in birds and other maniraptorans. Shorter feathers are preserved attached to the second finger.[6]
Scansoriopteryx had a non-perforated hip socket, which is more open in most, but not all, other dinosaurs. It also had a pubis (hip bone) which pointed forward, a primitive trait among theropods, and unlike some maniraptorans more closely related to birds, where the pubis points downward or backward.[6] The legs were short, and preserve small pebbly scales along the upper foot (metatarsus), as well as possible impressions of long feathers in the same area, possibly similar to the "hind wings" of Microraptor and other basal paravians.[6]
http://en.wikipedia.org/wiki/Epidexipteryx
A monophyletic Scansoriopterygidae was recovered by Godefroit et al. (2013); the authors found scansoriopterygids to be basalmost members of Paraves and the sister group to the clade containing Avialae and Deinonychosauria.[8]
ARCHAEOPTERYX
http://archosaurmusings.wordpress.com/2008/10/26/the-changing-legs-of-archaeopteryx/
One final point to add here is that the discovery of the wonderful 4-winged Microraptor gui seemed to come as complete shock to everyone. In hindsight of course it really should not have done so since Archaeopteryx clearly has long flight feathers on the legs.
Part of the program description
http://www.pbs.org/wgbh/nova/evolution/four-winged-dinosaur.html
The two markedly different reconstructions play into a long-running scientific controversy over the origin of flight in birds. For years the debate has been a standoff between two camps—those who believe dinosaurs were the ancestors of birds, and those who do not.
Believers in the dinosaur-bird connection have generally assumed that flight must have begun from the ground up, with fast-running dinosaurs that eventually got airborne as feathered arms evolved into wings, and running leaps evolved into powered flight.
Skeptics of the bird-dinosaur link say it would have been physically impossible for running dinosaurs to overcome gravity and get off the ground. It made more sense for flight to evolve from the trees down, with small, arboreal reptiles that glided from the treetops on their way to becoming full-fledged fliers. And that seemed to rule out dinosaurs, which presumably couldn't climb trees.
As seen in this program, the American Museum's Mark Norell is one of the proponents of the "birds-are-dinosaurs" hypothesis, which is the predominant view among most paleontologists, while Larry Martin of the University of Kansas speaks out for the minority view that birds descended from non-dinosaur tree dwellers.
Tantalizingly, Microraptor is the unexpected missing link that has reignited the debate and, with the help of NOVA's model and wind tunnel tests, just might settle the issue–or at the very least deepen our understanding of the long-ago era when the ancestors of birds first took to the air.
Possibilities
It seems that there may be three possibilities.
Ground up (dinosaurs)
Trees down (not dinosaurs)
A mishmash, with ground based dinosaurs developing feathers (for some other purpose than flight), then climbing trees, then using their feathers for the new purpose of gliding and eventually powered flight.
The problem with the mishmash, is that there is ZERO evidence for that.
To be even more precise, there is ZERO evidence for ground based dinosaurs climbing trees.
There is evidence of arboreal feathered creatures using their feathers for the purpose of gliding and powered flight. But there is no evidence that those arboreal feathered creatures had any connection to ground-based dinosaurs developing feathers (for some other purpose than flight) and then climbing trees.
Also, the mere presence of arboreal feathered creatures using their feathers for the purpose of gliding and powered flight, is no evidence of how they came to be.
Sunday, September 7, 2014
Propatagium (video)
Here you can see the extensive propatagium that it inherited from its pterosaur ancestor.
https://www.youtube.com/watch?v=YgNpkfKRQ-k
https://www.youtube.com/watch?v=YgNpkfKRQ-k
Wednesday, September 3, 2014
Oviraptors as secondarily flightless
Here is an accumulation of material on the idea that Oviraptors were secondarily flightless primitive birds. Being "secondarily" flightless means they evolved from earlier flying primitive birds.
Also see:
http://pterosaurnet.blogspot.ca/2015/06/oviraptor-propatagium.html
and
http://pterosaurnet.blogspot.com/2010/05/crests.html
http://en.wikipedia.org/wiki/Oviraptorosauria
http://en.wikipedia.org/wiki/Caudipteryx
http://agro.icm.edu.pl/agro/element/bwmeta1.element.agro-article-f38bf90a-7c6b-47fc-982a-30d0be44fd5e/c/app47-097.pdf
http://www.researchgate.net/publication/225452453_Pre-Archaeopteryx_coelurosaurian_dinosaurs_and_their_implications_for_understanding_avian_origins
http://www.nature.com/nature/journal/v455/n7216/full/nature07447.html
https://www.google.ca/url?sa=t&rct=j&q=&esrc=s&source=web&cd=3&cad=rja&uact=8&ved=0CDAQFjACahUKEwjigNb1vYHHAhWXG5IKHfNYBPQ&url=http%3A%2F%2Fdocencia.med.uchile.cl%2Fsmg%2Fpdf%2Fdinos2002.pdf&ei=XmC5VeLZDJe3yATzsZGgDw&usg=AFQjCNGlrsCF6bWr-XJndlp0iGVShn9LNg&sig2=IR_l7AMOalggmW_t5g74DQ&bvm=bv.99028883,d.aWw
This following is a counter to Jones et al (2000) but the problem with the following passage is that it uses the ambiguous phrase "non−avialan theropod". They do not rule out that oviraptors could be secondarily flightless members of Paraves.
http://www.app.pan.pl/archive/published/app50/app50-101.pdf
Oviraptors were either secondarily flightless avialae or secondarily flightless non-avialae paraves.
They descended from flying ancestors. They are not transitional between dinosaurs and paraves.
Here is a suggestion about the change that would accommodate the idea that oviraptors are secondarily flightless members of Eumaniraptora:
And here is how the so-called "non-paraves maniraptors" could be more correctly categorized as secondarily flightless birds:
Clade: Paraves
Subgroups
†Eosinopteryx
†Palaeopteryx
†Pneumatoraptor
†Scansoriopterygidae
†Yixianosaurus
Eumaniraptora
†Dromaeosauridae
†Troodontidae (including Jinfengopteryx)
Avialae
Oviraptorosauria
Alvarezsauria
Ornithomimosauria
Also see:
http://pterosaurnet.blogspot.ca/2015/06/oviraptor-propatagium.html
and
http://pterosaurnet.blogspot.com/2010/05/crests.html
http://en.wikipedia.org/wiki/Oviraptorosauria
Oviraptorosaurs Temporal range: Cretaceous, 130–66 Ma
Oviraptorosaurs, like deinonychosaurs, are so bird-like that several scientists consider them to be true birds, more advanced than Archaeopteryx. Gregory S. Paul has written extensively on this possibility, and Teresa Maryańska and colleagues published a technical paper detailing this idea in 2002.[5][15][16] Michael Benton, in his widely-respected text Vertebrate Paleontology, also included oviraptorosaurs as an order within the class Aves.[17] However, a number of researchers have disagreed with this classification, retaining oviraptorosaurs as non-avialan maniraptorans slightly more primitive than the deinonychosaurs.[18]
Analyses like those of Maryanska et al (2002) and Osmólska et al. (2004) suggest that they may represent primitive flightless birds.[5][6]
The consensus view, based on several cladistic analyses, is that Caudipteryx is a basal (primitive) member of the Oviraptoridae, and the oviraptorids are nonavian theropod dinosaurs.[9] Incisivosaurus is the only oviraptorid that is more primitive.[10]
Halszka Osmólska et al. (2004) ran a cladistic analysis that came to a different conclusion. They found that the most birdlike features of oviraptorids actually place the whole clade within Aves itself, meaning that Caudipteryx is both an oviraptorid and a bird. In their analysis, birds evolved from more primitive theropods, and one lineage of birds became flightless, re-evolved some primitive features, and gave rise to the oviraptorids. This analyis was persuasive enough to be included in paleontological textbooks like Benton's Vertebrate Paleontology (2005).[11] The view that Caudipteryx was secondarily flightless is also preferred by Gregory S. Paul,[12] Lü et al.,[13] and Maryańska et al.[14]
Others, such as Stephen Czerkas and Larry Martin have concluded that Caudipteryx is not a theropod dinosaur at all.[15] They believe that Caudipteryx, like all maniraptorans, is a flightless bird, and that birds evolved from non-dinosaurian archosaurs.[16]
http://agro.icm.edu.pl/agro/element/bwmeta1.element.agro-article-f38bf90a-7c6b-47fc-982a-30d0be44fd5e/c/app47-097.pdf
Avialan status for Oviraptorosauriahttp://www.ncbi.nlm.nih.gov/pubmed/19800747
TERESA MARYAŃSKA, HALSZKA OSMÓLSKA, and MIECZYSŁAW WOLSAN
Oviraptorosauria is a clade of Cretaceous theropod dinosaurs of uncertain affinities within Maniraptoriformes. All previous phylogenetic analyses placed oviraptorosaurs outside a close relationship to birds (Avialae), recognizing Dromaeosauridae or Troodontidae, or a clade containing these two taxa (Deinonychosauria), as sister taxon to birds. Here we present the results of a phylogenetic analysis using 195 characters scored for four outgroup and 13 maniraptoriform (ingroup) terminal taxa, including new data on oviraptorids. This analysis places Oviraptorosauria within Avialae, in a sister−group relationship with Confuciusornis. Archaeopteryx, Therizinosauria, Dromaeosauridae, and Ornithomimosauria are successively more distant outgroups to the Confuciusornis−oviraptorosaur clade. Avimimus and Caudipteryx are successively more closely related to Oviraptoroidea, which contains the sister taxa Caenagnathidae and Oviraptoridae. Within Oviraptoridae, “Oviraptor” mongoliensis and Oviraptor philoceratops are successively more closely related to the
Conchoraptor−Ingenia clade. Oviraptorosaurs are hypothesized to be secondarily flightless. Emended phylogenetic definitions are provided for Oviraptoridae, Caenagnathidae, Oviraptoroidea, Oviraptorosauria, Avialae, Eumaniraptora, Maniraptora, and Maniraptoriformes.
Recent studies by Varricchio et al. reveal that males cared for the eggs of troodontids and oviraptorids, so-called "non-avian theropods" of the Cretaceous, just as do those of most Paleognathic birds (ratites and tinamous) today. Further, the clutches of both groups have large relative volumes, and consist of many eggs of relatively large size. By comparison, clutch care by most extant birds is biparental and the clutches are of small relative volume, and consist of but few small eggs. Varricchio et al. propose that troodontids and oviraptorids were pre-avian and that paternal egg care preceded the origin of birds. On the contrary, unmentioned by them is that abundant paleontological evidence has led several workers to conclude that troodontids and oviraptorids were secondary flightless birds. This evidence ranges from bird-like bodies and bone designs, adapted for climbing, perching, gliding, and ultimately flight, to relatively large, highly developed brains, poor sense of smell, and their feeding habits.http://evolutionwiki.org/wiki/Paraves
Paraves, all maniraptorans closer to Neornithes than Oviraptor. Note that Sereno's excluding oviraptorosaurs from the paravian lineage is of questionable validity, as some of the most avian of all theropods, are oviraptorosaurs. Indeed, the clade may represent neoflightless [secondarily flightless] post-urvogels.
http://www.researchgate.net/publication/225452453_Pre-Archaeopteryx_coelurosaurian_dinosaurs_and_their_implications_for_understanding_avian_origins
the scansoriopterygids Epidendrosaurus [53] and Epidexipteryx[54] are more similar to basal birds, such as Jeholornis and Sapeornis [64,65], than to Archaeopteryx in many of their derived features, particularly in a number of derived cranial features. Surprisingly these cranial features are also seen in the oviraptorosaurs [54]. Together, the Jurassic maniraptorans suggest a monophyletic group composed of the scansoriopterygids,http://onlinelibrary.wiley.com/enhanced/doi/10.1111/j.1420-9101.2011.02349.x/
all other birds except Archaeopteryx, and probably also the oviraptorosaurs. This would represent a sister taxon to a monophyletic group containing the troodontids, the dromaeosaurids, and Archaeopteryx (Figure 1). Such a phylogenetic hypothesis would have significant implications for the reconstruction of the theropod-bird transition but it has yet to be tested by quantitative phylogenetic analysis.
Morphological gaps in the known fossil record separate only seven groups from the rest of Dinosauria. Those groups are Therizinosauroidea + Oviraptorosauria + Paraves, Tazoudasaurus + Eusauropoda, Ankylosauridae, Stegosauria, Neoceratopsia, basal Hadrosauriformes and Hadrosauridae.
http://www.nature.com/nature/journal/v455/n7216/full/nature07447.html
Here we report a new basal avialan, Epidexipteryx hui gen. et sp. nov., from the Middle to Late Jurassic of Inner Mongolia, China. This new species is characterized by an unexpected combination of characters seen in several different theropod groups, particularly the Oviraptorosauria. Phylogenetic analysis shows it to be the sister taxon to Epidendrosaurus4, 5, forming a new clade at the base of Avialae6.
https://www.google.ca/url?sa=t&rct=j&q=&esrc=s&source=web&cd=3&cad=rja&uact=8&ved=0CDAQFjACahUKEwjigNb1vYHHAhWXG5IKHfNYBPQ&url=http%3A%2F%2Fdocencia.med.uchile.cl%2Fsmg%2Fpdf%2Fdinos2002.pdf&ei=XmC5VeLZDJe3yATzsZGgDw&usg=AFQjCNGlrsCF6bWr-XJndlp0iGVShn9LNg&sig2=IR_l7AMOalggmW_t5g74DQ&bvm=bv.99028883,d.aWw
Jones et al (2000)
Caudipteryx has been described as a feathered dinosaur14,15 and
therefore would be expected to have had a dinosaurian mechanism
of cursoriality. However, relative total hindlimb proportions in
Caudipteryx contrast sharply with those in all other bipedal dinosaurs
and are indistinguishable from those in cursorial birds
(Fig. 1b). Accordingly, based on the tight linkage of hindlimb
proportions to cursorial mechanisms in bipedal archosaurs, we
suggest that Caudipteryx ran using a mechanism more similar to
that of modern cursorial birds than to typical dinosaurs. Significantly, lower leg (tibia + metatarsal) length in Caudipteryx is also the
same as the `effective hindlimb' length of cursorial birds, which is
equivalent to total hindlimb length in theropods (Fig. 1a).
This following is a counter to Jones et al (2000) but the problem with the following passage is that it uses the ambiguous phrase "non−avialan theropod". They do not rule out that oviraptors could be secondarily flightless members of Paraves.
http://www.app.pan.pl/archive/published/app50/app50-101.pdf
Caudipteryx zoui [an oviraptor] is a small enigmatic theropod known from the Early Cretaceous Yixian Formation of the People’s Republic of China. From the time of its initial description, this taxon has stimulated a great deal of ongoing debate regarding the phylogenetic relationship between non−avialan theropods and birds (Avialae) because it preserves structures that have been uncontroversially accepted as feathers (albeit aerodynamically unsuitable for flight). However, it has also been proposed that both the relative proportions of the hind limb bones (when compared with overall leg length), and the position of the center of mass in Caudipteryx are more similar to those seen in extant cusorial birds than they are to other non−avialan theropod dinosaurs. This conclusion has been used to imply that Caudipteryx may not have been correctly interpreted as a feathered non−avialan theropod, but instead that this taxon represents some kind of flightless bird. We review the evidence for this claim at the level of both the included fossil specimen data, and in terms of the validity of the results presented. There is no reason—phylogenetic, morphometric or otherwise—to conclude that Caudipteryx is anything other than a small non−avialan theropod dinosaur.NOTE:
Oviraptors were either secondarily flightless avialae or secondarily flightless non-avialae paraves.
They descended from flying ancestors. They are not transitional between dinosaurs and paraves.
Here is a suggestion about the change that would accommodate the idea that oviraptors are secondarily flightless members of Eumaniraptora:
Quote:
|
Clade: Paraves
Subgroups
†Eosinopteryx
†Palaeopteryx
†Pneumatoraptor
†Scansoriopterygidae
†Yixianosaurus
Eumaniraptora
†Dromaeosauridae
†Troodontidae (including Jinfengopteryx)
Avialae
Oviraptorosauria
Alvarezsauria
Ornithomimosauria
The other even more likely possibility is that some of these taxa have both flying AND secondarily flightless members.
https://bio.unc.edu/files/2011/04/FeducciaCzerkas2015.pdf
https://en.wikipedia.org/wiki/Scansoriopterygidae
https://books.google.ca/books?id=vtZFDb_iw40C&pg=PA165&lpg=PA165&dq=oviraptor+maryanska&source=bl&ots=FO917y8nqY&sig=H0mMsmdp0Diz6dVzCPmwLrPLwVo&hl=en&sa=X&ved=0CBwQ6AEwAGoVChMInbbrsLPHyAIViGseCh3qvQhH#v=onepage&q=oviraptor%20maryanska&f=false
https://www.insider.com/birds-that-look-like-dinosaurs-2019-2#with-4-inch-claws-and-a-tall-casque-on-its-head-the-cassowary-shares-physical-traits-with-prehistoric-creatures-2
https://bio.unc.edu/files/2011/04/FeducciaCzerkas2015.pdf
Feduccia and Czerkas
Considerable debate surrounds the numerous avian-like traits in core maniraptorans (oviraptorosaurs, troodontids, and dromaeosaurs), especially in the Chinese Early Cretaceous oviraptorosaur Caudipteryx, which preserves modern avian pennaceous primary remiges attached to the manus, as is the case in modern birds. Was Caudipteryx derived from earth-bound theropod dinosaurs, which is the predominant view among palaeontologists, or was it secondarily flightless, with volant avians or theropods as ancestors (the neoflightless hypothesis), which is another popular, but minority view. The discovery here of an aerodynamic propatagium in several specimens provides new evidence that Caudipteryx (and hence oviraptorosaurs) represent secondarily derived flightless ground dwellers, whether of theropod or avian affinity, and that their presence and radiation during the Cretaceous may have been a factor in the apparent scarcity of many other large flightless birds during that period.
https://en.wikipedia.org/wiki/Scansoriopterygidae
A subsequent phylogenetic analysis conducted by Agnolín and Novas (2011) recovered scansoriopterygids not as avialans, but as basal members of the clade Paraves remaining in unresolved polytomy with alvarezsaurids and the clade Eumaniraptora (containing avialans and deinonychosaurs).[8]Turner, Makovicky and Norell (2012) included only Epidexipteryx hui in their primary phylogenetic analysis, as a full-grown specimen of this species is known; regarding Scansoriopteryx/Epidendrosaurus, the authors were worried that including it in the primary analysis would be problematic, because it is only known from juvenile specimens, which "do not necessarily preserve all the adult morphology needed to accurately place a taxon phylogenetically" (Turner, Makovicky and Norell 2012, p. 89). Epidexipteryx was recovered as basal paravian that didn't belong to Eumaniraptora. The authors did note that its phylogenetic position is unstable; constraining Epidexipteryx hui as a basal avialan required two additional steps compared to the most parsimonious solution, while constraining it as a basal member of Oviraptorosauria required only one additional step. A separate exploratory analysis included Scansoriopteryx/Epidendrosaurus, which was recovered as a basal member of Avialae; the authors noted that it did not clade with Epidexipteryx, which stayed outside Eumaniraptora. Constraining the monophyly of Scansoriopterygidae required four additional steps and moved Epidexipteryx into Avialae.[9]A monophyletic Scansoriopterygidae was recovered by Godefroit et al. (2013); the authors found scansoriopterygids to be basalmost members of Paraves and the sister group to the clade containing Avialae and Deinonychosauria.[10] Agnolín and Novas (2013) recovered scansoriopterygids as non-paravian maniraptorans and the sister group to Oviraptorosauria.[11]http://digitallibrary.amnh.org/dspace/handle/2246/6352
Constraining Epidexipteryx as a basal oviraptorosaur requires only one additional step in our dataset (fig. 75). Three features support the inclusion of Epidexipteryx in Oviraptorosauria, which are caudal vertebrae without a transition point (char. 115.1), a dentary that has teeth only anteriorly (char. 220.1), and a first premaxillary tooth much larger than the succeeding teeth (char. 251.2). A tail without a transition point is unique to oviraptorosaurs and Epidexipteryx and premaxillary teeth greatly enlarged relative to other premaxillary teeth is unique to Incisivosaurus, Protarchaeopteryx, and Epidexipteryx.
Epidexipteryx resembles basal oviraptorosaurshttp://evolutionwiki.org/wiki/Velociraptor_a_Mesozoic_kiwi%3F_A_look_at_the_neoflightless_hypothesis
in several respects, particularly
in its cranial morphology. Zhang et al. (2008)
noted some of these, drawing attention to
the anteroposteriorly short but dorsoventrally
tall skull, the posterodorsally displaced
naris and anteroposteriorly long parietals.
Likewise, the highly procumbent anterior
dentition and the slightly downturned mandible
compares favorably to basal oviraptorosaurs
like Incisivosaurus, Caudipteryx, and
putatively Protarchaeopteryx.
Constraining Epidexipteryx as a basal oviraptorosaur
requires only one additional step in our dataset (fig. 75).”
https://books.google.ca/books?id=vtZFDb_iw40C&pg=PA165&lpg=PA165&dq=oviraptor+maryanska&source=bl&ots=FO917y8nqY&sig=H0mMsmdp0Diz6dVzCPmwLrPLwVo&hl=en&sa=X&ved=0CBwQ6AEwAGoVChMInbbrsLPHyAIViGseCh3qvQhH#v=onepage&q=oviraptor%20maryanska&f=false
https://www.insider.com/birds-that-look-like-dinosaurs-2019-2#with-4-inch-claws-and-a-tall-casque-on-its-head-the-cassowary-shares-physical-traits-with-prehistoric-creatures-2
Subscribe to:
Posts (Atom)