Pterosaur

ALTERNATIVES
https://en.wikipedia.org/wiki/Origin_of_birds#Digit_homology
The phalangeal count for archosaurs is 2-3-4-5-3
Thumbs Down
https://www.researchgate.net/publication/259253962_Thumbs_Down_A_MolecularMorphogenetic_Approach_to_Avian_Digit_Homology
We argue that limb bud cells that would normally form the digit II condensation proliferate toward a more anterior direction, into the space made physically available by the loss of digit I. This causes the presumptive digit II to leave the Shh activity zone. At the same time its cells do not express hoxD12 (and other posterior digit markers) any more, and therefore its transcriptome becomes characteristic of digit I. Its phalangeal number is reduced--whether due to weaker anterior FGF8 signaling, caused by lower Shh levels that would otherwise stabilize the expression, or because of different hox and downstream gene expression.
http://onlinelibrary.wiley.com/enhanced/doi/10.1002/jez.b.22545/#jezb22545-bib-0065
Thumbs down: a molecular-morphogenetic approach to avian digit homology
In the initial presentation of the concept, Kundrát et al. (2002) also suggested a mechanism that would be able to derive the Archaeopteryx phalangeal formula from the archosaur one. While the archosaur ground state is thought to be DI(2)–DII(3)–DIII(4)–DIV(5)–DV(3), Archaeopteryx could have DI(2)–DII(3)–DIII(4)–DV(0)–DV(0), but could also be interpreted as DI(0)–DII(2)–DIII(3)–DIV(4)–DV(0). Experiments with molecular signaling pathways in early limb development have shown that modulating interdigital bmp signaling (Dahn and Fallon, 2000) or blocking bmp with a dominant negative receptor (Zou and Niswander, 1996) is able to remove one phalanx from each digit, and therefore a mechanism like that could have caused the archosaur central digits to resemble the Archaeopteryx ones with regard to their phalangeal numbers.
Pyramid reduction hypothesis
The ‘pyramid reduction hypothesis’ assumes II-III-IV identities for neornithine manual digits and postulates the existence of a conservative five-digit pattern with a gradual, bilateral reduction of phalanges and metacarpals in avian evolution [9]. One proposed mechanism postulates that an elevation in peripheral BMPs, signaling factors that modulate cell survival and proliferation [60, 61], drove bilateral medial and lateral digital reduction [9]. This hypothesis is developmentally plausible, and is also consistent with the phalangeal reduction pattern seen in basal birds [9, 23]. However, it predicts that the direct avian ancestor had a five-fingered hand with dominant digits II, III, and IV [9], which is inconsistent with the digital reduction data from basal theropods (e.g., all known basal theropods, including ceratosaurs, have a vestigial digit IV) [5, 62, 63, 64]. In fact, the pyramid reduction hypothesis implies that either birds are not descended from theropod dinosaurs, or that some as yet to be discovered basal theropods were five-fingered with dominant digits II, III, and IV.
http://www.ncbi.nlm.nih.gov/pubmed/12210116?dopt=Abstract
We report herein that a pentadactyl developmental pattern is evident in early wing morphogenesis of Gallus (chicken) and Struthio (ostrich). Five avascular zones (spatially predestined locations of contiguous metacarpal and phalangeal aggregation) and four interdigital vascular spaces are established by the regression patterns of autopodial vasculature. Transient vestiges of the first and fifth metacarpals are confirmed histologically and histochemically. They lie within the preaxial-most and postaxial-most avascular zones, respectively.
These observations reveal conservative patterning of the avian hand and corroborate a II-III-IV metacarpal interpretation, argue for II-III-IV identity of ossified digits in birds, and favour a simple reduction rather than a homeotic [frame] shift in terms of the phenotype expressed by Hox genes in the phylogeny of the avian manus.
We suggest that gradual, bilateral reduction of phalanges and metacarpals, via apoptosis mediated by BMP, occurred during the evolution of birds (Pyramid Reduction Hypothesis). This is congruent with the establishment of a central wing axis that became co-opted for coordinated movements.
https://www.academia.edu/13081883/Pentadactyl_ground_state_of_the_avian_wing
The issue of the homology of bird fingers with those of pentadactyl amniotes has been a topic of contention for nearly 200 years. Data from the fossil record and phylogenetic systematics ascribe bird digit homologies to digits I, II, and III of pentadactyl amniotes while embryological evidence supports digital homologies of II, III, and IV. Using a molecular marker specific for condensation competent mesenchymal cells, we describe a pentadactyl arrangement of prechondrogenic digital anlagen in the wings of stage 29 chick embryos. Only the middle three anlagen develop into mature fingers. This pattern supports the hypothesis that bird fingers develop from digital anlagen II, III, and IV of pentadactylous amniotes. In addition, this result rejects a model assuming a shift in the primary axis in bird digit development and shows that a prechondrogenic digital anlage has been maintained in the bird lineage for at least 220 million years since the last known pentadactylous ancestor of the lineage. Such a vestige suggests that strong constraints are maintaining a pentadactyl ground state in amniotes.
https://angeldweb.wordpress.com/2016/07/04/problems-with-dinosaur-to-bird-evolution-the-iii-digits/
However, this view has remained inconsistent with the accepted early condensation patterns for digit progenitors, which show that the three hand digits of modern adult birds developed from the three middle positions. [14, 7, 9]. Furthermore, the assumption that a single frameshift mutation is capable of shifting Digits I, II III into II, III, IV has been shown to be unreliable given that such mutation reveals no adaptive advantage. Rather numerous frame shifts are required to even create an adaptive advantage, as demonstrated by Alan and other researchers. [15]Axis shift hypothesis
https://www.nature.com/articles/ncomms1437
Our results support the fossil record, indicating that birds evolved from theropod dinosaurs that lost digits 4 and 5 (Fig. 1a) and is consistent with the overlooked 'axis-shift' hypothesis that proposes the primary axis runs through the digit 3 position in bird wings19,20,21 (Figs 1f and g).
Alternative explanations for fingers
PRH, FS1, FS2, HOH, Thumbs Down, LSH.
And:
Very recently Xu and Mackem (2013) introduced a digit reduction scheme called the
lateral shift hypothesis (LSH). Like the TDH, it follows the II, III, IV
identification of tetanuran digits, and values the positional criteria over the compositional ones. Unlike the TDH, however, it argues
that a (partial) homeotic anterior shift took place and that digit IV
was completely re‐evolved, following in both points the FSH
Also see:
http://pterosaurnet.blogspot.ca/2014/07/pterosaur-fingers.html
https://en.wikipedia.org/wiki/Origin_of_birds#Digit_homology
ALTERNATIVES that are not credible
PROGENESIS
https://www.researchgate.net/profile/Tony_Thulborn/publication/43456034_Theropod_dinosaurs_progenesis_and_birds_Homology_of_digits_in_the_manus/links/567631d208ae0ad265c0e321.pdf
THUMBS DOWN
https://www.researchgate.net/publication/259253962_Thumbs_Down_A_MolecularMorphogenetic_Approach_to_Avian_Digit_Homology
https://frontiersinzoology.biomedcentral.com/articles/10.1186/1742-9994-11-33
https://www.researchgate.net/publication/259253962_Thumbs_Down_A_MolecularMorphogenetic_Approach_to_Avian_Digit_Homology
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3986427/
https://www.researchgate.net/publication/301913740_FANTASY_REALITY_A_Critique_of_Smith_'s_Bird_Origins
https://www.academia.edu/11078361/Pentadactyl_pattern_of_the_avian_wing_autopodium_and_pyramid_reduction_hypothesis
https://www.academia.edu/1486957/Transcriptomic_analysis_of_avian_digits_reveals_conserved_and_derived_digit_identities_in_birds
http://dml.cmnh.org/2000Dec/msg00446.html
Larry Febo
https://vargaslab.files.wordpress.com/2009/03/vargasfallon2005b3.pdf
https://pdfs.semanticscholar.org/c41f/be4e5fdc9b95780eefb7c396351290e43b23.pdf
https://www.academia.edu/13081883/Pentadactyl_ground_state_of_the_avian_wing
https://www.researchgate.net/publication/260714882_Tracing_the_Evolution_of_Avian_Wing_Digits
Xu and Mackem
http://people.eku.edu/ritchisong/554notes1.html

Also see:
https://pterosaurnet.blogspot.ca/2014/07/pterosaur-fingers.html
https://en.wikipedia.org/wiki/Origin_of_birds#Digit_homology
The phalangeal count for archosaurs is 2-3-4-5-3
ALTERNATIVES that are not credible
PROGENESIS
https://www.researchgate.net/profile/Tony_Thulborn/publication/43456034_Theropod_dinosaurs_progenesis_and_birds_Homology_of_digits_in_the_manus/links/567631d208ae0ad265c0e321.pdf
For instance, it is predictable that the loss of digit I from the manus should be simultaneous with loss of a single phalanx from one or more of the three remaining digits: as those losses are attributed to a single cause, the depletion of mesenehyme in the limb bud, they should not occur sequentially.
THUMBS DOWN
https://www.researchgate.net/publication/259253962_Thumbs_Down_A_MolecularMorphogenetic_Approach_to_Avian_Digit_Homology
We argue that limb bud cells that would normally form the digit II condensation proliferate toward a more anterior direction, into the space made physically available by the loss of digit I. This causes the presumptive digit II to leave the Shh activity zone. At the same time its cells do not express hoxD12 (and other posterior digit markers) any more, and therefore its transcriptome becomes characteristic of digit I.
Its phalangeal number is reduced--whether due to weaker anterior FGF8 signaling, caused by lower Shh levels that would otherwise stabilize the expression, or because of different hox and downstream gene expression.
Experiments with molecular signaling pathways in early limb development have shown that modulating interdigital bmp signaling (Dahn and Fallon, 2000) or blocking bmp with a dominant negative receptor (Zou and Niswander, '96) is able to remove one phalanx from each digit
A comparison of digits within each limb shows that the posterior forelimb digits are more strongly differentiated than the posterior hindlimb digits, and forelimb digit III exhibits a unique expression of Socs2.References
https://frontiersinzoology.biomedcentral.com/articles/10.1186/1742-9994-11-33
https://www.researchgate.net/publication/259253962_Thumbs_Down_A_MolecularMorphogenetic_Approach_to_Avian_Digit_Homology
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3986427/
https://www.researchgate.net/publication/301913740_FANTASY_REALITY_A_Critique_of_Smith_'s_Bird_Origins
https://www.academia.edu/11078361/Pentadactyl_pattern_of_the_avian_wing_autopodium_and_pyramid_reduction_hypothesis
https://www.academia.edu/1486957/Transcriptomic_analysis_of_avian_digits_reveals_conserved_and_derived_digit_identities_in_birds
http://dml.cmnh.org/2000Dec/msg00446.html
Larry Febo
https://vargaslab.files.wordpress.com/2009/03/vargasfallon2005b3.pdf
https://pdfs.semanticscholar.org/c41f/be4e5fdc9b95780eefb7c396351290e43b23.pdf
https://www.academia.edu/13081883/Pentadactyl_ground_state_of_the_avian_wing
https://www.researchgate.net/publication/260714882_Tracing_the_Evolution_of_Avian_Wing_Digits
Xu and Mackem
http://people.eku.edu/ritchisong/554notes1.html
Also see:
https://pterosaurnet.blogspot.ca/2014/07/pterosaur-fingers.html
http://onlinelibrary.wiley.com/doi/10.1002/jez.b.22545/full
http://www.researchgate.net/publication/259253962_Thumbs_down_a_molecular-morphogenetic_approach_to_avian_digit_homology
Avian forelimb digit homology remains one of the standard themes in comparative biology and EvoDevo research. In order to resolve the apparent contradictions between embryological and paleontological evidence a variety of hypotheses have been presented in recent years. The proposals range from excluding birds from the dinosaur clade, to assignments of homology by different criteria, or even assuming a hexadactyl tetrapod limb ground state. At present two approaches prevail: the frame shift hypothesis [FSH] and the pyramid reduction hypothesis [PRH]. While the former postulates a homeotic shift of digit identities, the latter argues for a gradual bilateral reduction of phalanges and digits.
We argue that limb bud cells that would normally form the digit II condensation proliferate toward a more anterior direction, into the space made physically available by the loss of digit I. This causes the presumptive digit II to leave the Shh activity zone. At the same time its cells do not express hoxD12 (and other posterior digit markers) any more, and therefore its transcriptome becomes characteristic of digit I. Its phalangeal number is reduced—whether due to weaker anterior FGF8 signaling, caused by lower Shh levels that would otherwise stabilize the expression, or because of different hox and downstream gene expression
Thus the change of the phalangeal formula (as in the PRH) is actually caused by the change of the transcriptome (as in the FSH), which in turn is directly caused by the loss of digit I (probably shh and hoxD mediated).
At the point when digit I is lost completely, the topological and morphogenetic effect causes the remaining digits to grow more toward anterior and therefore to adopt different phenotypic fates (e.g., Deinonychus).
The new approach presented here traces the origins of both theAlso:
FSH and the PRH to a common source: the loss of digit I. We
propose a morphogenetic‐molecular model in which the changingtranscriptome as well as the altered phalanx number in birds are
direct consequences of the reduction of the anterior‐most digit.Because this effect is thought to be triggered once digit I is lost, we
have dubbed it the thumbs down hypothesis (TDH)
https://www.researchgate.net/publication/261607127_New_developmental_evidence_supports_a_homeotic_frameshift_of_digit_identity_in_the_evolution_of_the_bird_wing
http://en.wikipedia.org/wiki/Bilateral_Digit_Reduction#Digit_homology
However, such frame shifts are rare in amniotes and—to be consistent with the theropod origin of birds—would have had to occur solely in the bird-theropod lineage forelimbs and not the hindlimbs (a condition unknown in any animal).[136] This is called Lateral Digit Reduction (LDR) versus Bilateral Digit Reduction (BDR) (see also Limusaurus[137])http://www.pnas.org/content/96/9/5111.full.pdf
Once this happened, we postulate a frame shift—a homeotic transformation—
in the developmental identity of the initial condensations:
Condensation CII developed into digit DI, CIII developed
into DII, and CIV developed into DIII (Fig. 6C), thus
conserving a functionally significant mature form within the
confines of a morphogenetic constraint.
We are not aware of any other case in which such a conflict between a developmental and a functional constraint in digit reduction existed.
http://onlinelibrary.wiley.com/doi/10.1002/jez.b.22545/full
http://www.researchgate.net/publication/259253962_Thumbs_down_a_molecular-morphogenetic_approach_to_avian_digit_homology
it argues that a (partial) homeotic anterior shift took place and that digit IV was completely re-evolved, following in both points the FSH.
http://www.cell.com/current-biology/pdf/S0960-9822(13)00512-5.pdf
Shh levels regulate digit pattern to shift their expression domains
for the developing hand and the re-emergence of a fully
functional digit in position 4
https://vargaslab.files.wordpress.com/2009/03/vargasfallon2005b3.pdf
Therefore, it must be understood clearly that the absence of a selective advantage for the developmental change in the wing does not imply that it could not have occurred.
From this point of view, the shift in digital identity could have occurred as a selectively neutral change (phenogenetic drift, according to Weiss and Fullerton, 2000)
http://frietsongalis.nl/wp-content/uploads/2009/06/Galis-et-al.-2005-J-Exp-Zool-B.pdf
In addition, a homeotic shift of digits 1, 2, 3 into digits 2, 3, 4 in theropods without further anatomical changes does not appear to lead to an adaptive advantage
GENERAL:
http://www.dinosauria.org/docs/oryctos/07_03_campbell.pdf
Even more revealing is the fact that in the Eichstätt specimen it can be seen that the ginglymoid joint at the end of Metacarpal II is rotated almost 90 degrees, such that in dorsal view it is clear that flexion/extension of Phalanx 1 of Digit II was primarily in the anterior/posterior plane
Significantly, an anteroposterior flexion/extension of Digit II, even with a sizable ventral component, would have made it the first avian alula, albeit a very primitive one.The basal paraves II digit is basically at right angles to the other two digits. It forms a "primitive alula". It has a quite different structure and function than the dinosaur 1st digit.
http://www.nature.com/nature/journal/v500/n7463/full/nature12336.html
Digit loss is defined as the complete loss of all phalanges and the
metapodial bone; it should be distinguished from digit reduction, in
which only phalanges are lost3. Digit loss can be adaptive. It reduces the
mass of the distal limb, and therefore its moment of inertia; this conserves
energy during running and flying12–14.
http://onlinelibrary.wiley.com/doi/10.1002/dvdy.22595/full


http://www.researchgate.net/publication/259253962_Thumbs_down_a_molecular-morphogenetic_approach_to_avian_digit_homology
https://www.academia.edu/1486957/Transcriptomic_analysis_of_avian_digits_reveals_conserved_and_derived_digit_identities_in_birds
To discover genes that specifically contribute to the second and third wing digit identities, we performed differential expression analysis of the mRNA-seq data between samples LFb and LFc. We found two genes, Tbx3 and Socs2, with high expression in sample LFc (Supplementary Fig. 9 and Fig. 3a). To our knowledge no studies have been published indicating a role for Socs2 in limb development. ISH confirms its strong expression in the third forelimb digit to the exclusion of all other digits in forelimb and hindlimb(Fig.3b–g). Recently it has been shown that the third forelimb digit has a unique mode of development in birds8. This, combined with our gene expression survey, supports the idea that the third wing digit has a unique derived identity in birds.
http://www.researchgate.net/publication/222677306_An_old_controversy_solved_bird_embryos_have_five_fingers
We are therefore left with several scenarios: (1) birdsAND
descending from archosaurs other than dinosaurs, which
cannot satisfactorily explain the many similarities
between birds and theropods; (2) the FSH, for which
there is as yet no adaptive significance that would
overcome the evolutionary constraint; and (3) birds
descending from theropods with digits II–IV, which is
the most parsimonious evolutionary transition scenario
but for which there is as yet no fossil evidence.
http://www2.gwu.edu/~newsctr/newscenter/research/dinosaur/clark.pdf
AND
http://blogs.discovermagazine.com/loom/2009/06/17/of-birds-and-thumbs/#.U-Okefnlp1Y
AND
http://genomebiology.com/content/pdf/gb-2011-12-10-130.pdf
AND
http://www.sciencedaily.com/releases/2011/09/110904140359.htm
AND
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3333764/
AND
http://www.nature.com/nature/journal/v459/n7249/abs/nature08124.html
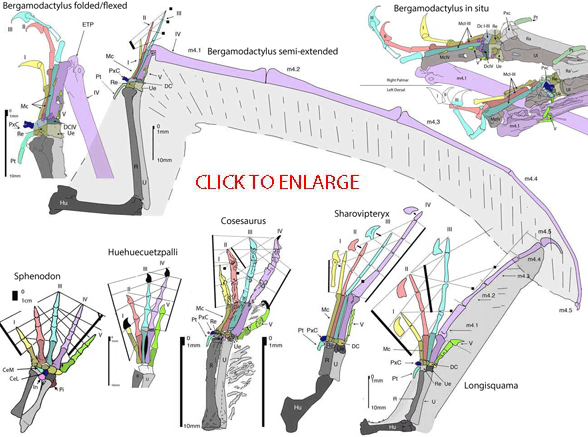
This may well be what the pterosaur hand looked like:
https://imaginosaurus.wordpress.com/tag/hands/

http://www.nature.com/nature/journal/v459/n7249/abs/nature08124.html
This may well be what the pterosaur hand looked like:
https://imaginosaurus.wordpress.com/tag/hands/
For a drawing of the pterosaur digits see page 33, Figure 4.10 (B):
https://books.google.ca/books?id=ND_PzHQuuLgC&pg=PA33&lpg=PA33&dq=witton+pterosaurs+page+35&source=bl&ots=a_VmiDU7A4&sig=98yv7sArfZ2-ym7iWAvzPq8-uk8&hl=en&sa=X&ved=0ahUKEwi6l4mN27PVAhVE6xQKHTgdAKAQ6AEITTAH#v=onepage&q=witton%20pterosaurs%20page%2035&f=false



{kind=link}