NESBITT (2011)
Nesbitt characters 212, 213, 214, 223, 230, 231 and 370.
212. Forelimb–hind limb, length ratio: (0) more than 0.55; (1) less than 0.55 (Gauthier,
1984; Sereno, 1991a; Juul, 1994; Benton, 1999).
Humerus + radius [forelimb] : Femur + tibia [hindlimb]
SEE REFERENCE IN DINOSAURIA (page 204):
Dromaeosaurid forelimbs are among the longest in theropods, the ratio of forelimb length to hindlimb length being about 65% in Velociraptor, 70% in Deinonychus and 80% in Sinornithosaurus.It seems that Nesbitt scored the Velociraptor incorrectly as (1).
Note that Velocirptor and Deinonychus are terrestrial, secondarily flightless members of Paraves.
213. Clavicles: (0) present and unfused; (1) fused into a furcula (modified from Gauthier, 1986; Sereno, 1991a; Benton, 1999; Benton and Walker, 2002). Clavicles are present in non-archosaurian archosauriforms and basal crocodylian-line archosaurs. Clavicles are not present in crocodylomorphs (e.g., Hesperosuchus ‘‘agilis,’’ CM 29894; Protosuchus richardsoni, AMNH FR 3024) and, therefore, they are scored as inapplicable. Like the interclavicle, the clavicles of the pterosaur Eudimorphodon are separate ossifications in a small specimen and incorporated into the sternum (Wild, 1993). All other pterosaurs seem to lack distinct ossifications of the clavicles. Within Dinosauria, clavicles are present, but do not contact in some ornithischians (e.g., Psittacosaurus) and are unossified in others (Butler et al., 2008a). The clavicles of some nonsauropod sauropodomorphs (e.g., Massospondylus) may contact each other at the midline, but do not fuse (Yates and Vasconcelos, 2005). A furcula (fused clavicles) is present in nearly all theropods known from complete
 skeletons including Coelophysis bauri (AMNH FR 30647; Rinehart et al., 2007; Nesbitt et al., 2009d) and Allosaurus fragilis (UUVP 6102; Chure and Madsen, 1996). This character has been employed by various datasets exploring theropod relationships (e.g., Norell et al., 2001; Clarke, 2004).
skeletons including Coelophysis bauri (AMNH FR 30647; Rinehart et al., 2007; Nesbitt et al., 2009d) and Allosaurus fragilis (UUVP 6102; Chure and Madsen, 1996). This character has been employed by various datasets exploring theropod relationships (e.g., Norell et al., 2001; Clarke, 2004).Pterosaurs have clavicles and an interclavicle that are fused into the sternum.
214. Interclavicle: (0) present; (1) absent
(fig. 30) (Gauthier, 1986; Sereno, 1991a; Juul,
1994; Benton, 1999).
The interclavicle is present in archosauriforms
plesiomorphically (Sereno, 1991a) and
persists through Pseudosuchia. In Pterosauria,
an interclavicle appears to be present
in young individuals of Eudimorphodon
(MCSNB 8950), but fuse to the pectoral
elements in larger individuals (Wild, 1993). A
distinct interclavicle is not present in all other
pterosaurs. Ornithischians and saurischians
lack an interclavicle. However, the pectoral
girdles in the successive sister taxa to
Dinosauria (Silesaurus, Marasuchus, Lagerpeton)
do not have the pectoral region
completely preserved. As a result, the optimization
of this character within Dinosauromorpha
is not clear.
SEE REFERENCE IN DINOSAURIA (page 204)
Also from Nesbitt et al (2009) (page 872):
The evolutionary transformation of the furcula
from separate clavicles is nicely illustrated in
archosaurs and their close relatives. Euparkeria
capensis and early pseudosuchians retain both the
interclavicle and clavicles. The interclavicle is lost
at the dinosaur node or at an unknown node
among early dinosauromorphs. From there, the origin
of the furcula is well understood with the
addition of the work on basal sauropodomorphs of
Yates and Vasconcelos (2005) and the discovery of
furculae in early coelophysoid theropods (Tykoski
et al., 2002; Rinehart et al., 2007). Some ornithischians
have two small clavicles that do not
contact each other (Osborn, 1924a; Brown and
Schlaikjer, 1940; Sternberg, 1951; Chinnery and
Weishampel, 1998). In contrast, saurischians
retain clavicles with the clavicles contacting at the
midline.
Birds and pterosaurs have an interclavicle. Dinosaurs do not.
230. Humerus, apex of deltopectoral crest situated at a point corresponding to: (0) less than 30% down the length of the humerus; (1) more than 30% down the length of the humerus (fig. 31) (modified from Bakker and Galton, 1974; Benton, 1990a; Juul, 1994; Novas, 1996; Benton, 1999).
Langer and Benton (2006) thoroughly discussed the distribution of the character states of this character and find that state (1) is restricted to dinosaurs within Archosauria. Here, I follow the conclusions and scorings of Langer and Benton (2006).
The pterosaur deltopectoral crest is like birds. The dinosaur deltopectoral crest is not like birds. (See details below).
231. Humerus, length: (0) longer than or
subequal to 0.6 of the length of the femur; (1)
shorter than 0.6 of the length of the femur
(modified from Novas, 1996; Langer and
Benton, 2006).
Langer and Benton (2006) thoroughly
discussed the distribution of the character
states and find that state (1) is restricted to
Herrerasaurus (PVSJ 373), Eoraptor (PVSJ
512), and neotheropods.
Humerus length compared to femur length is similar for pterosaurs to birds but different than dinosaurs.
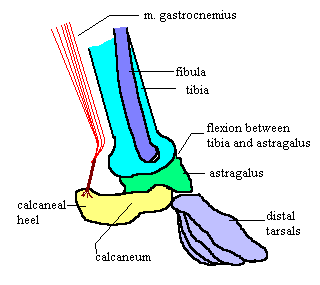
370. Astragalus-calcaneum, articulation:
(0) free; (1) coossified (fig. 46) (Sereno and
Arcucci, 1994a; Irmis et al., 2007a).
In most archosauriforms, save avians and
close relatives, the astragalus and calcaneum
are separate elements. In pterosaurs (e.g.,
Dimorphodon, YPM 9182), Lagerpeton (PVL
4619), and Dromomeron romeri (GR 223), the
astragalus and calcaneum are coossified.
Among basal dinosaurs, the proximal tarsals
are coossified in Heterodontosaurus (SAMPK-
1332) and coelophysoids (Rowe and
Gauthier, 1990; Tykoski, 2005b).

More info on the deltopectoral crest.
http://palaeos.com/vertebrates/glossary/glossaryD.html
We see that the dinosaur furcula is not like the Paravian furcula.
That supports the conclusion that they are not related.
Also see (page 77)
http://books.google.ca/books?id=8QRKV7eSqmIC&pg=PA77&lpg=PA77&dq=oviraptor+furcula&source=bl&ots=fqQ1dMcCAm&sig=Mh6VWAuDyk92AmCENy_FFdCRvn0&hl=en&sa=X&ei=g-t4U96CENKdyASEzYDYAw&ved=0CGAQ6AEwCQ#v=onepage&q=oviraptor%20furcula&f=false
THE EARLY EVOLUTION OF ARCHOSAURS:
RELATIONSHIPS AND THE ORIGIN OF
MAJOR CLADES 92011)
from separate clavicles is nicely illustrated in
archosaurs and their close relatives. Euparkeria
capensis and early pseudosuchians retain both the
interclavicle and clavicles. The interclavicle is lost
at the dinosaur node or at an unknown node
among early dinosauromorphs. From there, the origin
of the furcula is well understood with the
addition of the work
on basal sauropodomorphs ofYates and Vasconcelos (2005) and the discovery of
furculae in early coelophysoid theropods (Tykoski
et al., 2002; Rinehart et al., 2007). Some ornithischians
have two small clavicles that do not
contact each other (Osborn, 1924a; Brown and
Schlaikjer, 1940; Sternberg, 1951; Chinnery and
Weishampel, 1998). In contrast, saurischians
retain clavicles with the clavicles contacting at the
midline.
Birds and pterosaurs have an interclavicle. Dinosaurs do not.
223. Coracoid, postglenoid process: (0) short; (1) elongate and expanded posteriorly only
The significance of this is that a longer coracoid allows the bird scapula (attached to the coracoid) to be positioned high enough on the body (horizontally), to allow flapping (by the wings being raised high enough up).
The significance of this is that a longer coracoid allows the bird scapula (attached to the coracoid) to be positioned high enough on the body (horizontally), to allow flapping (by the wings being raised high enough up).
Birds are like pterosaurs and not like dinosaurs.
Langer and Benton (2006) thoroughly discussed the distribution of the character states of this character and find that state (1) is restricted to dinosaurs within Archosauria. Here, I follow the conclusions and scorings of Langer and Benton (2006).
The pterosaur deltopectoral crest is like birds. The dinosaur deltopectoral crest is not like birds. (See details below).
231. Humerus, length: (0) longer than or
subequal to 0.6 of the length of the femur; (1)
shorter than 0.6 of the length of the femur
(modified from Novas, 1996; Langer and
Benton, 2006).
Langer and Benton (2006) thoroughly
discussed the distribution of the character
states and find that state (1) is restricted to
Herrerasaurus (PVSJ 373), Eoraptor (PVSJ
512), and neotheropods.
Humerus length compared to femur length is similar for pterosaurs to birds but different than dinosaurs.
370. Astragalus-calcaneum, articulation:
(0) free; (1) coossified (fig. 46) (Sereno and
Arcucci, 1994a; Irmis et al., 2007a).
In most archosauriforms, save avians and
close relatives, the astragalus and calcaneum
are separate elements. In pterosaurs (e.g.,
Dimorphodon, YPM 9182), Lagerpeton (PVL
4619), and Dromomeron romeri (GR 223), the
astragalus and calcaneum are coossified.
Among basal dinosaurs, the proximal tarsals
are coossified in Heterodontosaurus (SAMPK-
1332) and coelophysoids (Rowe and
Gauthier, 1990; Tykoski, 2005b).
More info on the deltopectoral crest.
http://palaeos.com/vertebrates/glossary/glossaryD.html
Deltopectoral crest: a longitudinal ridge or crest on the (proximal) humerus. See figure. Cursorial forms typically do nothave a large crest. It is typically an important attachment point for adductors, rather than retractors.http://pterosaurheresies.wordpress.com/category/pterosaur-evolution
"Both tiny birds and tiny pterosaurs dispensed with their long stiff tail. In birds it became a pygostyle. In pterosaurs the long stiff tail became a reduced, string-like tail with bead-like verts. Note the similarities in the pectoral girdles. Both could stand with their toes beneath their shoulder glenoids. Both had retroverted pedal digits but of two distinct designs. The anterior ilium of both taxa supported large thigh muscles. A large deltopectoral crest supported large flight adductors anchored to the sternum."
Deltopectoral crest: a longitudinal ridge or crest on the (proximal) humerus. Cursorial forms typically do not have a large crest. It is typically an important attachment point for adductors, rather than retractors.
http://en.wikipedia.org/wiki/Evolution_of_dinosaurs

http://www.scielo.br/scielo.php?script=sci_arttext&pid=S0001-37652009000400017&lng=es&nrm=iso&tlng=es
Pterosaur humerus:

See page 117.
It appears that the pterosaur deltopectoral crest differed between the long-bony-tailed pterosaurs and the short-bony-tailed pterosaurs.
In both cases they were confined to close to the proximal end of the humerus. But the deltopectoral crest in the long-bony-tailed pterosaurs was much smaller.
The deltopectoral crest in the primitive birds was close to the proximal end and relatively small.
In contrast the deltopectoral crest in dinosaurs was elongate. It was not confined to the proximal end of the humerus.
http://courses.washington.edu/chordate/453photos/skeleton_photos/bird-humeri.jpg

Notice the bird deltopectoral crest is confined to the proximal end as in pterosaurs.
http://en.wikipedia.org/wiki/Dinosaur#Distinguishing_anatomical_features
Dinosaurs evolved within a single lineage of archosaurs 232-234 Ma (million years ago) in the Ladinian age, the latter part of the middle Triassic. Dinosauria is a well-supported clade, present in 98% of bootstraps. It is diagnosed by many features including loss of the postfrontal on the skull and an ELONGATE deltopectoral crest on the humerus.[1]
The Dinosauria: Second Edition:
The humerus is slender and twisted. It has a caudally deflected proximal end and a moderately developed deltopectoral crest that is restricted to the PROXIMAL third of the humerus in Deinonychus and to the PROXIMAL quarter in Velociraptor.
The deltopectoral crest on mosthttps://pterosaurheresies.files.wordpress.com/2015/03/aurornis-humerus.jpgdinosaurs, including Archaeopteryx (Fig. 1), is prominent and extends down a quarter of the humerus. We don’t see this in Scansioropteryx (Fig. 3). But then again, we don’t see this in Aurornis (Fig. 4) either.That doesn’t delete them from the theropod clade because every other aspect of their anatomy says: theropod!
http://www.scielo.br/scielo.php?script=sci_arttext&pid=S0001-37652009000400017&lng=es&nrm=iso&tlng=es
Pterosaur humerus:

Description: The specimen MB.R. 2828 (Fig. 2; Table I) has a slightly dorso-ventral compression, showing several fractures. It possesses the characteristic saddle-shaped proximal articular head, common to the pterosaur humeri. The deltopectoral crest is well developed and inclined proximoventrally. It is tongue-shaped with a rounded distal end. In lateral view the proximal margin of the deltopectoral crest is markedly concave, while the distal margin is straight. There is an elongated ridge on the medial side of the crest running from the distal to the proximal edge, and is likely an attachment of a flight muscle (m. pectoralis, see Bennett 2003). There is an elongated concavity on the medial side, close to the distal margin, whose function is unknown. The ulnar crest is blunt and slightly crushed laterally. The dorsal margin shows a well developed pneumatic foramen, located close to the lateral side.https://books.google.ca/books?id=8CKYxcylOycC&pg=PA117&lpg=PA117&dq=rhamphorhynchus+deltopectoral+crest&source=bl&ots=SqlXdvwI83&sig=2eO8UZBiUTQPDTye_KvTu59iYQw&hl=en&sa=X&ei=MQuQVd_BKpGGNsCtifgC&ved=0CB0Q6AEwAA#v=onepage&q=rhamphorhynchus%20deltopectoral%20crest&f=false
See page 117.
It appears that the pterosaur deltopectoral crest differed between the long-bony-tailed pterosaurs and the short-bony-tailed pterosaurs.
In both cases they were confined to close to the proximal end of the humerus. But the deltopectoral crest in the long-bony-tailed pterosaurs was much smaller.
The deltopectoral crest in the primitive birds was close to the proximal end and relatively small.
In contrast the deltopectoral crest in dinosaurs was elongate. It was not confined to the proximal end of the humerus.
http://courses.washington.edu/chordate/453photos/skeleton_photos/bird-humeri.jpg
Miscellaneous bird humeri (proximal heads to the left). The top 2 face anteriorly & the bottom 3 face posteriorly.

Notice the bird deltopectoral crest is confined to the proximal end as in pterosaurs.
http://en.wikipedia.org/wiki/Dinosaur#Distinguishing_anatomical_features
apex of deltopectoral crest (a projection on which the deltopectoral muscles attach) located at or more than 30% down the length of the humerus (upper arm bone)
Notice the dinosaur deltopectoral crest is not confined to the proximal end as in birds and pterosaurs. It is elongate.
NESBITT (2009)
| 11. Furcula shape in anterior view: asymmetrical (0) or symmetrical/nearly symmetrical (1). The furculae of most nonparavian theropods are markedly asymmetrical (e.g., Allosaurus, Citipati). The furculae of paravians, with the exception of Buitreraptor, are nearly symmetrical. It is unclear if the asymmetry of the furcula of Buitreraptor was the result of taphonomy or represents morphological asymmetry. |
That supports the conclusion that they are not related.
Also see (page 77)
http://books.google.ca/books?id=8QRKV7eSqmIC&pg=PA77&lpg=PA77&dq=oviraptor+furcula&source=bl&ots=fqQ1dMcCAm&sig=Mh6VWAuDyk92AmCENy_FFdCRvn0&hl=en&sa=X&ei=g-t4U96CENKdyASEzYDYAw&ved=0CGAQ6AEwCQ#v=onepage&q=oviraptor%20furcula&f=false
THE EARLY EVOLUTION OF ARCHOSAURS:
RELATIONSHIPS AND THE ORIGIN OF
MAJOR CLADES 92011)
The character states supporting pterosaurs
as members of Archosauria and Ornithodira
are not restricted to character states related
to locomotion as suggested by Bennett
(1996). As demonstrated in the list above,
the character states cover features present all
over the body, not just in the hind limb.
Furthermore, it is difficult to argue that the
restricted number of tarsals, the size of the
distal tarsals, and the shape of the proximal
tarsals in pterosaurs would be convergent
with those of dinosauromorphs based on
function alone (Sereno 1991a). In summary,
Pterosauromorpha is well supported as the
sister taxon to Dinosauromorpha.
No comments:
Post a Comment