I am willing to pay $500 for a cladistic analysis on the relationship of birds to pterosaurs..
If you are interested, please submit a comment including a reference link to a site where I can evaluate your qualifications.
Thursday, December 26, 2013
Monday, December 16, 2013
Respiratory cycle of a bird (aspiration pump)
Birds have and pterosaurs had the same breathing system.
BIRDS
http://www.peteducation.com/article.cfm?c=15+1829&aid=2721

http://svpow.com/2013/12/11/unidirectional-airflow-in-the-lungs-of-birds-crocs-and-now-monitor-lizards/

PTEROSAURS
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2637988/
The following references confirm that birds have an "avian aspiration pump" but dinosaurs did not.
http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0003303
Evidence for Avian Intrathoracic Air Sacs in a New Predatory Dinosaur from Argentina
BIRDS
http://www.peteducation.com/article.cfm?c=15+1829&aid=2721

- During the first inspiration, the air travels through the nostrils, also called nares, of a bird, which are located at the junction between the top of the upper beak and the head. The fleshy tissue that surrounds them, in some birds, is called the cere. As in mammals, air moves through the nostrils into the nasal cavity. From there it passes through the larynx and into the trachea. Air moves through the trachea to the syrinx, which is located at the point just before the trachea divides in two. It passes through the syrinx and then the air stream is divided in two as the trachea divides. The air does not go directly to the lung, but instead travels to the caudal (posterior) air sacs. A small amount of air will pass through the caudal air sacs to the lung.
- During the first expiration, the air is moved from the posterior air sacs through the ventrobronchi and dorsobronchi into the lungs. The bronchi continue to divide into smaller diameter air capillaries. Blood capillaries flow through the air capillaries and this is where the oxygen and carbon dioxide are exchanged.
- When the bird inspires the second time, the air moves to the cranial air sacs.
- On the second expiration, the air moves out of the cranial air sacs, through the syrinx into the trachea, through the larynx, and finally through the nasal cavity and out of the nostrils.
http://svpow.com/2013/12/11/unidirectional-airflow-in-the-lungs-of-birds-crocs-and-now-monitor-lizards/

PTEROSAURS
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2637988/
Respiratory Evolution Facilitated the Origin of Pterosaur Flight and Aerial Gigantism (2009)
(Leon P. A. M. Claessens1*, Patrick M. O'Connor2, David M. Unwin3)
In this report we present various lines of skeletal evidence that indicate that pterosaurs had a highly effective flow-through respiratory system, capable of sustaining powered flight, predating the appearance of an analogous breathing system in birds by approximately seventy million years.
The skeletal breathing pump of pterosaurs, including the vertebral and sternal ribs, sternum, gastralia and prepubes, likely formed a highly integrated functional complex. The persistence of the basic components of this system in all pterosaur clades suggests that our inferences related to ventilatory mechanics, and primarily based upon Rhamphorhynchus and Pteranodon, can be safely assumed to have generally applied to the group.
"The aspiration pump of pterosaurs maximised trunk expansion in the ventrocaudal region, while at the same time limiting the degrees of freedom of movement of the trunk in other directions. This provided greater control over the location, amount and timing of trunk expansion, thereby enabling precisely-timed localized generation of pressure gradients within the pulmonary system, a trait that is also present in living birds where it is of paramount importance for the generation of air flow patterns in the lungs [27], [36].
The following references confirm that birds have an "avian aspiration pump" but dinosaurs did not.
http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0003303
Evidence for Avian Intrathoracic Air Sacs in a New Predatory Dinosaur from Argentina
Paul C. Sereno et al
Research on the gastral cuirass in archosaurs led to the suggestion that it may have functioned as an accessory aspiration pump in nonavian dinosaurs [81]–[83]. Although Claessens drew attention to the relationship between the gastral cuirass and abdominal air sacs, he concluded that “it appears impossible to ascertain exactly when lung diverticula stretching throughout the whole body cavity or unidirectional airflow originated” [83:102]. Later a “caudal origin model” for air sacs and flow-through lung ventilation (either uni- or bidirectional) was proposed [33: 255] based on (1) the presence of abdominal air sacs (inferred from posterior dorsal and sacral pneumaticity), (2) a dynamic gastral cuirass, and (3) vertebrocostal articulations in the posterior ribcage that allow greater excursion during aspiration (inferred from the more horizontal arrangement of posterior rib articulations). An independent study of rib morphology, in contrast, concluded that nonavian dinosaurs were characterized by an “anteriorly ventilated bellows lung”[84: 47].
Tuesday, July 9, 2013
Questions?
If anyone has questions, I will do my best to answer them.
Please use a name other than "anonymous". It need not be your actual name.
Please use a name other than "anonymous". It need not be your actual name.
Friday, June 28, 2013
Further focus (2)
Continuing with the hypothesis that:
Long tailed pterosaurs developed into Paraves which then developed into short tailed Pygostylia (Avialae).

Time-calibrated phylogeny showing the temporal range of the main pterosaur clades; basal clades in red, pterodactyloids in blue; known ranges of clades indicated by solid bar, inferred ‘ghost’ range by coloured line; footprint symbols indicate approximate age of principal pterosaur track sites based on Lockley et al. (2008); stratigraphic units and age in millions of years based on Gradstein et al. (2005). 1, Preondactylus; 2, Dimorphodontidae; 3, Anurognathidae; 4, Campylognathoididae; 5, Scaphognathinae; 6, Rham- phorhynchinae; 7, Darwinopterus; 8, Boreopterus; 9, Istiodactylidae; 10, Ornithocheiridae; 11, Pteranodon; 12, Nyctosauridae; 13, Pterodactylus; 14, Cycnorhamphus; 15, Ctenochasmatinae; 16, Gnathosaurinae; 17, Germanodactylus; 18, Dsungaripteridae; 19, Lonchodectes; 20, Tapejaridae; 21, Chaoyangopteridae; 22, Thalassodromidae; 23, Azhdarchidae. Abbreviations: M, Mono- fenestrata; P, Pterodactyloidea; T, Pterosauria; ca, caudal vertebral series; cv, cervical vertebral series; mc, metacarpus; na, nasoantorbital fenestra; r, rib; sk, skull; v, fifth pedal digit.
Darwinopterus is in there, too—it’s the small purple box numbered “7″.

Thursday, June 27, 2013
Further focus (1)
Let us examine a more detailed hypothesis that:
Long-tailed pterosaurs developed into basal Paraves which later developed into short-tailed Pygostylia .
http://en.wikipedia.org/wiki/Deinonychosauria
The teeth of deinonychosaurs were curved and serrated, but not blade-like except in some advanced species such as Dromaeosaurus albertensis. The serrations on the front edge of deinonychosaur teeth were very small and fine, while the back edge had serrations which were very large and hooked.[3] Deinonychosaurs generally had long, winged forelimbs, though these were smaller in some troodontids. The wings usually bore three large, flexible claws.[3]http://en.wikipedia.org/wiki/Pygostylia
Pygostylia is a group of birds which includes Confuciusornis and all of the more derived birds; the Ornithothoraces. [2]The pygostylians fall into two distinct groups with regard to the pygostyle. The Ornithothoraces have a ploughshare-shaped end, while the more primitive members have a longer, rod-shaped pygostyle.http://en.wikipedia.org/wiki/Avialae
Avialans diversified into a wide variety of forms during the Cretaceous Period.[25] Many groups retained primitive characteristics, such as clawed wings and teeth, though the latter were lost independently in a number of bird groups, including modern birds (Neornithes). While the earliest forms, such as Archaeopteryx and Jeholornis, retained the long bony tails of their ancestors,[25] the tails of more advanced birds were shortened with the advent of the pygostyle bone in the clade Pygostylia. In the late Cretaceous, around 95 million years ago, the ancestor of all modern birds also evolved better olfactory senses.[26]http://en.wikipedia.org/wiki/Ornithurae
Ornithurae (meaning "bird tails" in Greek) is the name of a natural group which includes the common ancestor of Ichthyornis, Hesperornis, and all modern birds as well as all other descendants of that common ancestor.
Thursday, February 21, 2013
Significant separation
http://en.wikipedia.org/wiki/Coelurosauria
The two most significant separations between subgroups are those between the Paraves andThe reason that there is a "significant separation" between Paraves and coelurosaur dinosaurs is because they are not related.othercoelurosaurs and between the paravian clades Avialae and Deinonychosauria.
Tuesday, February 19, 2013
Paraves existed at the same time as the earliest coelurosaur dinosaurs
Note that Paraves (Eumaniraptora) existed at the same time as the earliest coelurosaur dinosaurs. Paraves were primitive birds with pennaceous feathers. Paraves could not have descended from coelurosaur dinosaurs.
(Click to enlarge).

Note Eumaniraptora in the chart above. It is on the far right. You can see that it extends back to the time of the very earliest found coelurosaur dinosaurs.
(Click to enlarge).
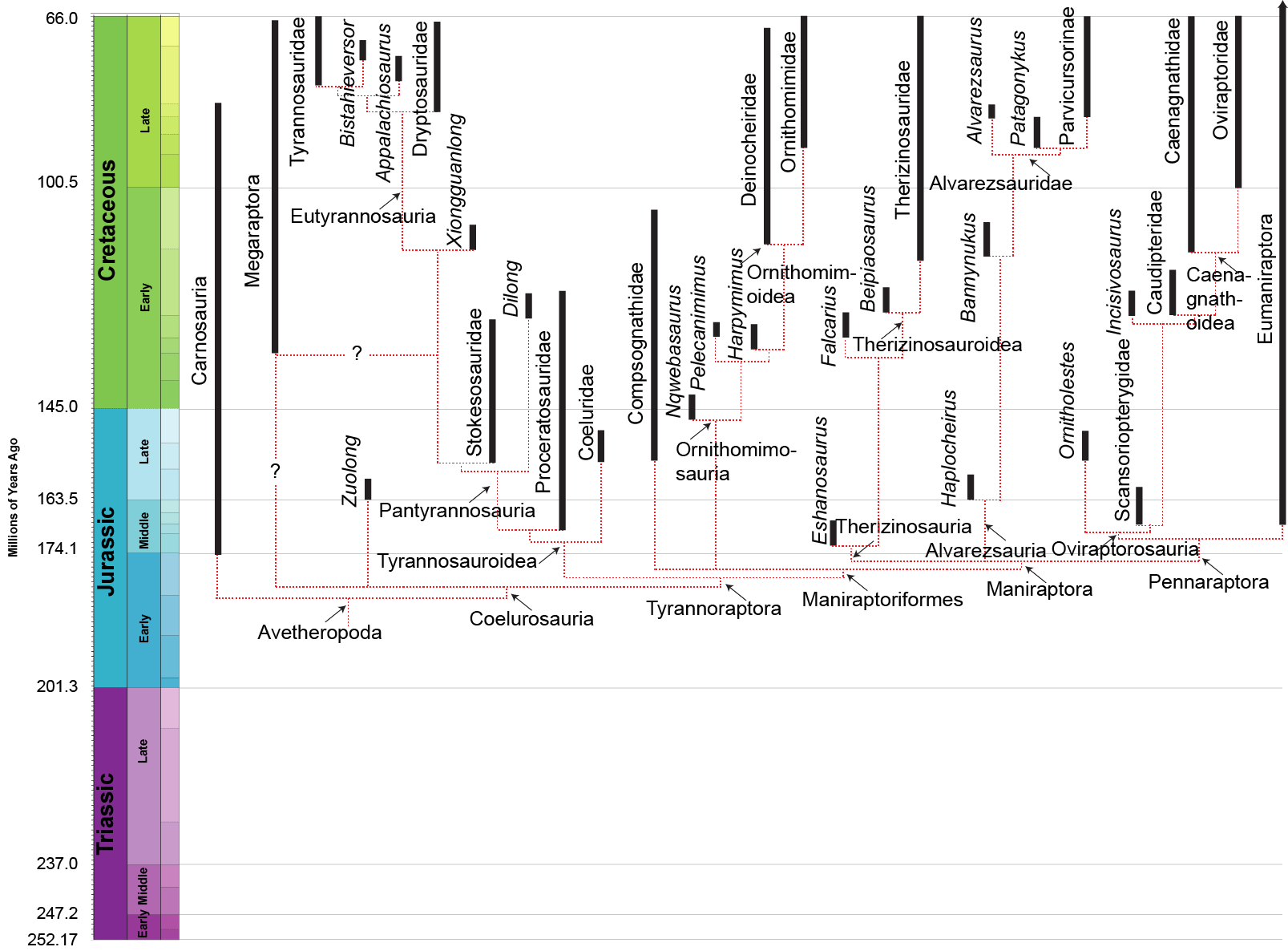
Note Eumaniraptora in the chart above. It is on the far right. You can see that it extends back to the time of the very earliest found coelurosaur dinosaurs.
Friday, February 15, 2013
Pterosaurs taken as birds
Here are pterosaurs that have been taken as birds. This again shows that pterosaurs and primitive birds are very similar.
http://en.wikipedia.org/wiki/Palaeocursornis
http://en.wikipedia.org/wiki/Piksi
http://journals2.scholarsportal.info/details.xqy?uri=/12809659/v34i0004/883_sropbvaprtab.xml
http://en.wikipedia.org/wiki/Palaeocursornis
Palaeocursornis is a monotypic genus of pterosaurs. The only known species, P. corneti, was described in 1984 based on a single bone (MTCO-P 1637) interpreted as the distal part of a left femur, found in Early Cretaceous (Berriasian rocks (dating to around 143 mya) from a mine at Cornet near Oradea in northwestern Romania. It was initially assumed to be a flightless paleognathe bird, possibly a ratite, and later as a more primitive ornithuromorph or non-avialan theropod (Benton et al., 1997). However, re-evaluation of the specimen suggested that it was not a femur at all, but the upper arm bone (humerus) of a pterodactyloid pterosaur similar to Azhdarcho.[1]
http://en.wikipedia.org/wiki/Piksi
Piksi is a genus of pterosaurs containing the single species Piksi barbarulna (meaning "strange elbowed big bird ", from Blackfoot piksi, "big bird" or, specifically, "chicken" and Latin barbarus "strange, outlandish" + ulna, elbow[1]). It lived roughly 75 million years ago in what is now Montana, USA. Known from parts of a right wing – the humerus, ulna and radius bones – the only specimens found so far are housed in the Museum of the Rockies(collection number MOR 1113). The genus Piksi is monotypic at present.
The bones are fragmentary and represent roughly the elbow area. Comparing the fossils' size to the wing bones of other ground birds, P. barbarulna seems to have been about as large as a Common Pheasant, i.e. some 15 in (35–40 cm) long excluding tail, and with a wingspan of perhaps 30 in (80 cm) or somewhat less. It would thus have weighed maybe 1 – 2 pounds (some 500 g – 1 kg).[2]The original description of the fossils found its affinities unresolvable except that it was probably an ornithothoracine bird. Agnolin and Varricchio (2012) reinterpreted Piksi barbarulna as a pterosaur rather than a bird, most likely a member of Ornithocheiroidea.[3]Original article::
http://journals2.scholarsportal.info/details.xqy?uri=/12809659/v34i0004/883_sropbvaprtab.xml
Varricchio (2002 ) described some forelimb bones from the Late Cretaceous (Campanian) Two Medicine Formation, Glacier County, Montana (USA), as the holotype of Piksi barbarulna, a supposed ornithothoracine bird. However reevaluation of Piksi Varricchio, 2002 instead recognizes this genus as belonging to Pterosauria Kaup, 1834 and not Aves Linnaeus, 1758. Piksi exhibits the following derived humeral traits of pterosaurs: 1) very large ectepicondyle; 2) large trochlea; 3) with a deep, wide and poorly deliminated brachial depression that is proximodistally extended; 4) a wide and deep olecranal fossa not marked dorsally by a ridge; and 5) lacking a distal depression of the groove for the m. humerotricipitalis. Moreover, the putative Early Cretaceous birds Eurolimnornis Jurcsák & Kessler, 1986 and Palaeocursornis Jurcsák & Kessler, 1986 , based on distal humeri, are also regarded as pterosaurs. The record of Piksi constitutes an important addition to the Latest Cretaceous pterosaurian record.
Sunday, February 3, 2013
Flight Stroke
The basal Paraves used the same flight stroke as Pterosaurs which is sufficient for flapping flight. The following references correctly show that neither pterosaurs nor basal paraves used their m. supracoracoideus as a pulley. But they are wrong in asserting that that precluded flapping flight.
PTEROSAURS
http://press.princeton.edu/witton/sampler-pterosaurs.pdf
http://biology.kenyon.edu/courses/biol241/bird%20flight%202003%20Chatterjee_Sankar.pdf
Basal paraves inherited their characteristics from their pterosaur ancestor and evolved feathers to replace the skin membranes.
They continued to use the same flight stroke from their pterosaur ancestor.
Basal Paraves are feathered pterosaurs.
The evidence strongly supports the transition from pterosaur to basal paraves, with Scansoriopterygidae (or a taxon much like it) as being transitional between pterosaur and basal paraves
We have seen evidence that Scansoripteryx is one of the most basal members of paraves.
It could splay its hind limbs like pterosaurs. It used the same muscles as pterosaurs for flight.
Scansoriopterygidae (or a taxon much like it) is an excellent candidate for transitional between pterosaurs and later basal paraves.
SPLAYED HINDLIMBS
There is a reference to the splayed posture of Scansoriopterids here (page 154):
Riddle of the Feathered Dragons
http://books.google.ca/books?id=SihlpQTlVdAC&pg=PA154&lpg=PA154&dq=parasagittal+stance+for+Archaeopteryx.&source=bl&ots=jTl0YCn6be&sig=B9WKpEcJJ8Xr3U4_haeHeZFUToE&hl=en&sa=X&ei=xHMYVMWbAsGOyAT3loGQDg&ved=0CDwQ6AEwAw#v=snippet&q=in%20a%20splayed%20posture&f=false
There is a reference to the splayed posture of Archaeopteryx here (page 399):
The Origin and Evolution of Birds
http://books.google.ca/books?id=8QRKV7eSqmIC&pg=PA399&lpg=PA399&dq=archaeopteryx+femur+angle&source=bl&ots=fqR1hR9GAi&sig=7grokjyLDiND0WyvJNaAJC8e0Ko&hl=en&sa=X&ei=p1UYVJW0H5OtyATf5YCYCA&ved=0CB0Q6AEwAA#v=onepage&q=archaeopteryx%20femur%20angle&f=false
http://dml.cmnh.org/2010Feb/msg00281.html
The feathered pterosaur hindwings replace the function of the uropatagium of their pterosaur ancestor just as the wings replace the function of the patagium.
And of course the feathered pterosaur propatagium is simply the pterosaur propatagium.
http://www.aou.org/auk/content/130/1/0001-0013.pdf
Feduccia
Alexander et al
"Tent" model:
http://icb.oxfordjournals.org/content/early/2011/09/21/icb.icr112.full
Also chicks can have a problem with legs that are splayed too much.
https://www.google.ca/webhp?sourceid=chrome-instant&rlz=1C1SKPL_enCA423CA423&ion=1&espv=2&ie=UTF-8#q=birds%20splay%20their%20legs
http://en.wikipedia.org/wiki/Microraptor
http://www.app.pan.pl/archive/published/app59/app20111109.pdf
Pterosaur pelvis
http://www.pnas.org/content/107/7/2733.full.pdf+html
John Ruben
http://www.pnas.org/content/106/13/5002.abstract?ijkey=b80c6cec04b5f05bcbf67870ea44df19a403ed62&keytype2=tf_ipsecsha
Hesperonychus
http://www.pnas.org/content/107/40/E155.full
Counter-argument (Brougham and Brusatte)
http://www.pnas.org/content/107/40/E156.full
Reply to Brougham and Brusatte
FOR REFERENCE:
MODERN BIRD
http://en.wikipedia.org/wiki/Bird_anatomy


PTEROSAURS
http://press.princeton.edu/witton/sampler-pterosaurs.pdf
At one stage, it was thought that the flight muscles ofhttp://eurekamag.com/research/019/478/morphological-evolution-pectoral-girdle-pterosaurs-myology-role.php (2003)
pterosaurs were very birdlike, with the arm lifted by
a muscle, m. supracoracoideus, anchoring on the sternum
rather than the shoulders. In birds, this muscle
arcs over the glenoid to attach on the dorsal surface
of the humerus, elevating the wing with a pulley-like
system (e.g., Kripp 1943; Padian 1983a; Wellnhofer
1991a). Detailed reconstruction of the proximal arm
musculature of pterosaurs shows that this is not
the case, however, and that the [pterosaur] arm was more likely
lifted by large muscles anchored on the scapula and
back, and lowered by those attached to the sternum
and coracoid (fig. 5.8; Bennett 2003a). Unlike [modern] birds,
where two vastly expanded muscles are mainly used
to power flight, it appears that pterosaurs used several
muscle groups to form their flapping strokes.
The musculature of the pectoral region of representative rhamphorhynchoid (Campylognathoides) and large pterodactyloid (Anhanguera) pterosaurs was reconstructed in order to examine the function of various muscles and the functional consequences of the evolution of the advanced pectoral girdle of large pterodactyloids. The reconstructions suggest that m. supracoracoideus was not an elevator of the wing, but instead depressed and flexed the humerus. m. latissimus dorsi, m, teres major, m. deltoides scapularis, and m. scapulohumeralis anterior were wing elevators.BASAL PARAVES
http://biology.kenyon.edu/courses/biol241/bird%20flight%202003%20Chatterjee_Sankar.pdf
Its lack of a supracoracoideus (SC)
pulley, the primary elevator of the wing, would prevent
Archaeopteryx from executing humeral rotation on the
glenoid during the upstroke,
For example, Archaeopteryx could nothttp://www.ncbi.nlm.nih.gov/pmc/articles/PMC1780066/
position its wing high in an upstroke position, since it
lacked a modern avian supracoracoideus (SC) pulley, the
primary elevator of the wing (Poore et al. 1997).
Anatomical evidence indicates that Microraptor was not capable of ground or running takeoff, because it lacked the supracoracoideus pulley to elevate the wings.http://en.wikipedia.org/wiki/Confuciusornis
The first of these regarded problems to attain a steep flight path due to a limited wing amplitude. In the interpretation of Senter (2006) of the position of the shoulder joint, a normal upstroke would be impossible precluding flapping flight entirely. Less radical is the assessment that due to the lack of a keeled sternum and a high acrocoracoid, the [Confuciusornis] Musculus pectoralis minor could not serve as a M. supracoracoideus lifting the humerus via a tendon running through a foramen triosseum. This, coupled with a limited upstroke caused by a lateral position of the shoulder joint, would have made it difficult to gain altitude.http://jeb.biologists.org/content/200/23/2987.full.pdf
The lack of a morphologically derived SC in Late
Jurassic and Early Cretaceous birds precluded a high velocity
recovery stroke which undoubtedly limited
powered flight in these forms. Subsequent evolution of the
derived SC capable of imparting a large rotational force to
the humerus about its longitudinal axis was an important
step in the evolution of the wing upstroke and in the ability
to supinate (circumflex) the manus in early upstroke, a
movement fundamental to reducing air resistance during
the recovery stroke.
The highly derived morphology of the SC,Pterosaurs and basal paraves flapped their wings in the same way.
a characteristic of modern birds capable of powered flight, was
not present in Archaeopteryx (Ostrom, 1976a,b; Wellnhofer,
1988, 1993), nor is there firm evidence for its presence in
recently described Mesozoic species (Chiappe, 1995; Sanz et
al. 1996).
| Scansoriopterygidae are basal paraves with flight feathers on the arms and legs. Scansoriopterygidae (or a group very much like it) is the ancestor of the later Paraves such as Microraptor, Archaeopteryx etc. Rhamphorhynchidae (or a group very much like it) is the ancestor of Scansoriopterygidae. Rhamphorhynchidae pycnofibres are homologous to Scansoriopterygidae feathers. The Rhamphorhynchidae acetabulum is homologous to the Scansoriopterygidae acetabulum. The Rhamphorhynchidae caudal rods are homologous to the Scansoriopterygidae caudal rods. The Rhamphorhynchidae long bony tail is homologous to the Scansoriopterygidae long bony tail. The Scansoriopterygidae outermost digit is transitional between Rhamphorhynchidae and later Paraves. |
They continued to use the same flight stroke from their pterosaur ancestor.
Basal Paraves are feathered pterosaurs.
The evidence strongly supports the transition from pterosaur to basal paraves, with Scansoriopterygidae (or a taxon much like it) as being transitional between pterosaur and basal paraves
We have seen evidence that Scansoripteryx is one of the most basal members of paraves.
It could splay its hind limbs like pterosaurs. It used the same muscles as pterosaurs for flight.
Scansoriopterygidae (or a taxon much like it) is an excellent candidate for transitional between pterosaurs and later basal paraves.
SPLAYED HINDLIMBS
There is a reference to the splayed posture of Scansoriopterids here (page 154):
Riddle of the Feathered Dragons
http://books.google.ca/books?id=SihlpQTlVdAC&pg=PA154&lpg=PA154&dq=parasagittal+stance+for+Archaeopteryx.&source=bl&ots=jTl0YCn6be&sig=B9WKpEcJJ8Xr3U4_haeHeZFUToE&hl=en&sa=X&ei=xHMYVMWbAsGOyAT3loGQDg&ved=0CDwQ6AEwAw#v=snippet&q=in%20a%20splayed%20posture&f=false
There is a reference to the splayed posture of Archaeopteryx here (page 399):
The Origin and Evolution of Birds
http://books.google.ca/books?id=8QRKV7eSqmIC&pg=PA399&lpg=PA399&dq=archaeopteryx+femur+angle&source=bl&ots=fqR1hR9GAi&sig=7grokjyLDiND0WyvJNaAJC8e0Ko&hl=en&sa=X&ei=p1UYVJW0H5OtyATf5YCYCA&ved=0CB0Q6AEwAA#v=onepage&q=archaeopteryx%20femur%20angle&f=false
http://dml.cmnh.org/2010Feb/msg00281.html
A dynamic, modular flight system (as in birds) required a chaotic, unstable system to operate in, and modulating the body to compensate. Nonexistent or gently controlled air flow is not a good way to determine performance of a glide path, especially since NO living glider today maintains an absolutely fixed physical posture during its trajectory, and this goes triple for parachuting animals.
In their conclusion, Alexander et al. state: "Obviously, the living animalIn fact flying pterosaurs did precede them.
was capable of active control, but we suggest that the tandem wing
configuration may have been advantageous because it requires less active
stabilizing ability."
I am not quite sure how one can develop a system of
unstable flight control from a passively stable system. Abilities to develop
control over the wing and perform a dynamic powered operation seem to require a
powered operation to precede it
The feathered pterosaur hindwings replace the function of the uropatagium of their pterosaur ancestor just as the wings replace the function of the patagium.
And of course the feathered pterosaur propatagium is simply the pterosaur propatagium.
http://www.aou.org/auk/content/130/1/0001-0013.pdf
Feduccia
Microraptors have been reconstructed in two distinctive models, the four-winged gliding model with sprawled hindlimb wings, by which it was originally described in Nature (Xu et al. 2003), and a dinosaurian bipedal model, or biplane model, by which it is reconstructed with the hindlimbs held beneath the body, incapable of sprawling, in other words, like a tiny T. rex. The problem,of course, is that there is absolutely no reason the hindlimbs could not have been sprawled, as is the case in flying squirrels (Glaucomys spp.), flying lemurs (Dermoptera), etc., and even falling cats. Too, the sprawled model performs superiorly inwind-tunnel experiments (Alexander et al. 2010), most specimensare preserved with a sprawled posture, and the wingclaws are adapted for trunk climbing (Burnham et al. 2011). In addition, it would be difficult to imagine how selection could produce elongate, asymmetric hindlimb flight remiges by the most current paleontological reconstructions, in which the hindlimbs are held in flight beneath the body in obligate bipedal fashion, with elongate hindlimb wing feathers trailing behind, simply slicing through the air (Balter 2012)http://www.pnas.org/content/107/7/2972.full.pdf+html
Alexander et al
Fossils of the remarkable dromaeosaurid Microraptor gui and relatives clearly show well-developed flight feathers on the hind limbs as well as the front limbs. No modern vertebrate has hind limbsfunctioning as independent, fully developed wings; so, lacking a living example, little agreement exists on the functional morphologyor likely flight configuration of the hindwing. Using a detailed reconstruction based on the actual skeleton of one individual, cast in the round, we developed light-weight, three-dimensional physical models and performed glide tests with anatomically reasonable hindwing configurations. Models were tested with hindwings abducted and extended laterally, as well as with a previously described biplane configuration. Although the hip joint requiresthe hindwing to have at least 20° of negative dihedral (anhedral),all configurations were quite stable gliders. Glide angles rangedfrom 3° to 21° with a mean estimated equilibrium angle of 13.7°,giving a lift to drag ratio of 4.1:1 and a lift coefficient of 0.64. The abducted hindwing model’s equilibrium glide speed corresponds to a glide speed in the living animal of 10.6m·s−1. Although the biplane model glided almost as well as the other models, it was structurally deficient and required an unlikely weight distribution (very heavy head) for stable gliding. Our model with laterally abducted hindwings represents a biologically and aerodynamically reasonable configuration for this four-winged gliding animal. M. gui’s feathered hindwings, although effective for gliding, would have seriously hampered terrestrial locomotion.
Primitively, early archosaurs are sprawling, with the legs set
laterally and elevated at around 75° (6), a preadapted posture for
gliding. Modern birds normally have the thigh elevated and
sprawled to the side in different degrees; for example, it is nearly
perpendicular to the midline in loons and grebes (7). This variation
shows that the degree of splaying needed to use the
hindlegs in gliding is not unusual when compared with that in
modern birds. The absence of an antitrochanter and a supraacetabular
acetabular shelf (SAC) in the eumaniraptorans, including dromaeosaurids,
would make elevation and splaying of the legs even
easier (8). Air pressure could have provided most of the force
needed to elevate the leg into a gliding position similar to that in
gliding mammals. This simple positioning was originally assumed
for the four-winged Microraptor gui (5); but later, workers hoping
to recover an upright posture proposed arrangements of the
hindlimbs that would have required complicated systems of locks
and muscles to hold the leg in an only partially elevated position,
e.g., the “biplane” model (9). New anatomical information based
on the discovery of several hundred specimens similar to the
four-winged glider M. gui (and related taxa) has produced converging
lines of evidence demonstrating that the original
describers of M. gui (5) were correct in their interpretation of the
flight posture. We postulate, based on examination of this new
material, that M. gui was capable of abducting the hind limbs at
least 65–70° to achieve a gliding posture.
"Tent" model:
http://icb.oxfordjournals.org/content/early/2011/09/21/icb.icr112.full
Also chicks can have a problem with legs that are splayed too much.
https://www.google.ca/webhp?sourceid=chrome-instant&rlz=1C1SKPL_enCA423CA423&ion=1&espv=2&ie=UTF-8#q=birds%20splay%20their%20legs
http://en.wikipedia.org/wiki/Microraptor
Some paleontologists have doubted the biplane hypothesis, and have proposed other configurations. A 2010 study by Alexander et al. described the construction of a lightweight three-dimensional physical model used to perform glide tests. Using several hind leg configurations for the model, they found that the biplane model, while not unreasonable, was structurally deficient and needed a heavy-headed weight distribution for stable gliding, which they deemed unlikely. The study indicated that a laterally abducted hindwing structure represented the most biologically and aerodynamically consistent configuration for Microraptor.[3] A further analysis by Brougham and Brusatte, however, concluded that Alexander's model reconstruction was not consistent with all of the available data on Microraptor and argued that the study was insufficient for determining a likely flight pattern for Microraptor. Brougham and Brusatte criticized the anatomy of the model used by Alexander and his team, noting that the hip anatomy was not consistent with other dromaeosaurs. In most dromaeosaurids, features of the hip bone prevent the legs from splaying horizontally; instead, they are locked in a vertical position below the body. Alexander's team used a specimen of Microraptor which was crushed flat to make their model, which Brougham and Brusatte argued did not reflect its actual anatomy.[15] Later in 2010, Alexander's team responded to these criticisms, noting that the related dromaeosaur Hesperonychus, which is known from complete hip bones preserved in three dimensions, also shows hip sockets directed partially upward, possibly allowing the legs to splay more than in other dromaeosaurs.[16]Related references:
http://www.app.pan.pl/archive/published/app59/app20111109.pdf
Pterosaur pelvis
http://www.pnas.org/content/107/7/2733.full.pdf+html
John Ruben
http://www.pnas.org/content/106/13/5002.abstract?ijkey=b80c6cec04b5f05bcbf67870ea44df19a403ed62&keytype2=tf_ipsecsha
Hesperonychus
http://www.pnas.org/content/107/40/E155.full
Counter-argument (Brougham and Brusatte)
http://www.pnas.org/content/107/40/E156.full
Reply to Brougham and Brusatte
FOR REFERENCE:
MODERN BIRD
http://en.wikipedia.org/wiki/Bird_anatomy
The supracoracoideus works using a pulley like system to lift the wing while the pectorals provide the powerful downstroke


Monday, January 28, 2013
Pelvic Bones Summary
Here is a summary of the material about pterosaur, dinosaur and basal paraves pelvic bones.
See also this link
http://pterosaurnet.blogspot.ca/2010/05/acetabulum.html


http://pterosaurnet.blogspot.ca/2010/05/pubic-bones.html
http://pterosaurnet.blogspot.ca/2012/12/no-connection-between-dinosaurs-and.html
http://www.aou.org/auk/content/130/1/0001-0013.pdf
The transition to partially open acetabulum (from fully closed) would have taken place in the transition from pterosaur to basal paraves.
GENERAL:
http://books.google.ca/books?id=idta6AVV-tIC&pg=PA10&lpg=PA10&dq=Unwin+%281988%29+,+pterosaurs+have+an+imperforate+acetabulum,&source=bl&ots=2E_W7T0hQs&sig=sEqzrX-ZuACLWeCHl23YaV9z9gc&hl=en&ei=2TvoS6W2B4bGlQfmxfnXAw&sa=X&oi=book_result&ct=result&resnum=3&ved=0CCAQ6AEwAg#v=onepage&q=Unwin%20(1988)%20%2C%20pterosaurs%20have%20an%20imperforate%20acetabulum%2C&f=false
http://pterosaurheresies.wordpress.com/2011/10/14/pterosaur-femur-time/
Two pterodactylid pterosaurs:

http://www.lusofossils.com/verte.html
Dinosaur:

But notice the Anchiornis femur (d):
http://www.ivpp.ac.cn/qt/papers/201403/P020140314379333425828.pdf
http://scienceblogs.com/tetrapodzoology/wp-content/blogs.dir/471/files/2012/04/i-3f9364c23349a5c25c429724b628ee11-Anchiornis_details_Jan_2009.jpg
 Figure 4 IVPP V 14378 selected elements. (a) Anteriormost caudal vertebrae in ventral view; (b) left carpal region; (c) right ilium in lateral view; (d) left femur in posteromedial view; (e) left pes in dorsolateral view. Abbreviations: bs, brevis fossa; ?dc3, ?distal carpal 3; fh, femoral head; MTII-IV, metatarsals II-IV; mw, medial wall; pst, supra-trochanteric process; ra, radius; rle, radiale; sac, superacetabular crest; ?‘se’, ?‘semilunate carpal; ul, ulna; ?ule, ?ulnare. Scale bar = 3 mm for (a)―(d) and 1 mm for (e).
Figure 4 IVPP V 14378 selected elements. (a) Anteriormost caudal vertebrae in ventral view; (b) left carpal region; (c) right ilium in lateral view; (d) left femur in posteromedial view; (e) left pes in dorsolateral view. Abbreviations: bs, brevis fossa; ?dc3, ?distal carpal 3; fh, femoral head; MTII-IV, metatarsals II-IV; mw, medial wall; pst, supra-trochanteric process; ra, radius; rle, radiale; sac, superacetabular crest; ?‘se’, ?‘semilunate carpal; ul, ulna; ?ule, ?ulnare. Scale bar = 3 mm for (a)―(d) and 1 mm for (e).
Bambiraptor:
http://kuscholarworks.ku.edu/dspace/...ries.013op.pdf

Hesperonychus:
http://www.pnas.org/content/106/13/5002.full

MODERN BIRD
http://www.innerbird.com/pelvic_girdle/pelvic_girdle.html

Note the pterosaur prepubis:
http://pterosaurheresies.wordpress.com/2011/11/06/whats-with-that-deep-prepubis/
http://archosaurmusings.wordpress.com/2009/12/24/back-to-that-pterosaur-sacrum-pelvis
The Auk 124(3):789–805, 2007
http://en.wikipedia.org/wiki/Pterosaur
https://www.app.pan.pl/archive/published/app59/app20111109.pdf
https://en.wikipedia.org/wiki/Sacrum
https://en.wikipedia.org/wiki/Synsacrum
http://www.prehistoriclife.xyz/history/mesozoic-birds.html
http://www.reinhold-necker.de/seite14.html
http://bio.sunyorange.edu/updated2/pl%20new/55%20first%20birds.htm
https://static-content.springer.com/image/art%3A10.1186%2F2041-9139-5-25/MediaObjects/13227_2014_Article_146_Fig3_HTML.jpg

http://www.ivpp.ac.cn/qt/papers/201403/P020140314389417822583.pdf
http://www.app.pan.pl/article/item/app20120093.html
W. Scott Persons, IV, Philip J. Currie, and Mark A. Norell
http://brantworks.com/resources/Allosaurus/ANTS_kit/ANTS-Allosaurus-Frank-Galef-detail-spine-BrantWorks.jpg

http://www.pterosaur.co.uk/PDB2012/P/index.htm
See also this link
http://pterosaurnet.blogspot.ca/2010/05/acetabulum.html
PELVIC BONES
http://icb.oxfordjournals.org/cgi/content/full/40/1/87#SEC3
http://pterosaur.net/fossils_coloborhynchus_pelvis.php"Several aspects of the pelvic girdle suggest that pterosaurs were specialized for pelvic aspiration during flight. As in birds, the three pelvic bones of pterosaurs were solidly fused into a single unit (Fig. 9), and an increased number of dorsal vertebrae were incorporated into the sacrum (3 to 5 in Rhamphorhynchusand as many as 10 in Pteranodon) (Wellnhofer, 1978http://www.livescience.com/3399-north-american-dinosaur-smaller-housecat.html, 1987
"The claws were thought to come from juveniles — they were just so small. But when we studied the pelvis, we found the hip bones were fused, which would only have happened once the animal was fully grown," Longrich said. "Until now, the smallest carnivorous dinosaurs we have seen in North America have been about the size of a wolf. Judging by the amount of material that was collected, we believe animals the size of Hesperonychus [a dromaeosaurid] must have been quite common on the landscape."

The pelvis of the orinithocheiroid Coloborhynchus.http://pterosaur.net/fossils/coloborhynchus_pelvis_2.jpg
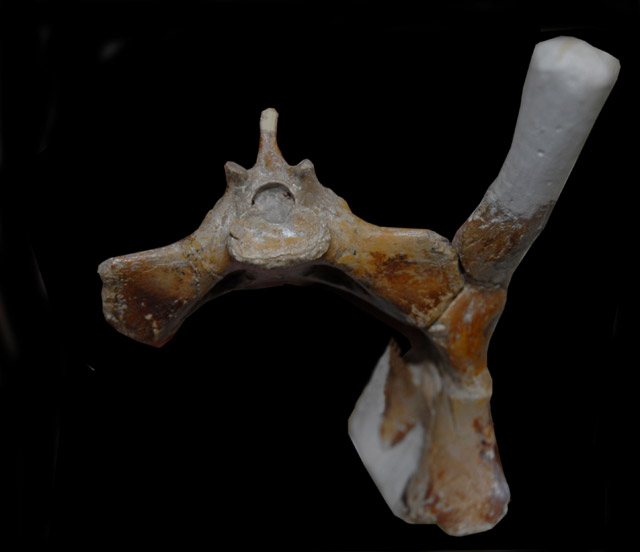
http://pterosaurnet.blogspot.ca/2010/05/pubic-bones.html
http://pterosaurnet.blogspot.ca/2012/12/no-connection-between-dinosaurs-and.html
ACETABULUM/FEMUR ARTICULATION
SUMMARY:http://www.aou.org/auk/content/130/1/0001-0013.pdf
A partially closed acetabulum is seen in basal archosaurs and is characteristic of the scansoriopterids and Jurassic feathered forms such as Anchiornis initially described as near Aves by Xu et al (2009)
GENERAL:
See page 10
The hip joint of pterosaurs is more mobile and profoundly different from that of theropods but is reminiscent of that of mammals, especially of humans, allowing a wide range of adduction and abduction in the vertical plane.
The femoral component [of the Anhanguera pterosaur femur] is a well defined spherical head which is distinctly separated from the shaft by a narrow non-articular neck at an obtuse angle of 160 degrees. The head forms a ball and socket joint with the close-fitting, shallow and imperforate acetabulum.
In theropods [dinosaurs], the femoral component is cylindrical without any distinctive head and neck. It projects medially at a right angle [90 degrees] from the shaft and fits into a perforated acetabulum of up to 1.5 times its diameter. As a result, the hip joint is stable and fully congruent during parasagittal motion, permitting a wide range of flexion and extension but very little abduction and adduction.
http://pterosaurheresies.wordpress.com/2011/10/14/pterosaur-femur-time/
Two pterodactylid pterosaurs:
http://www.lusofossils.com/verte.html
Dinosaur:

But notice the Anchiornis femur (d):
http://www.ivpp.ac.cn/qt/papers/201403/P020140314379333425828.pdf
http://scienceblogs.com/tetrapodzoology/wp-content/blogs.dir/471/files/2012/04/i-3f9364c23349a5c25c429724b628ee11-Anchiornis_details_Jan_2009.jpg
Anchiornis:

Bambiraptor:
http://kuscholarworks.ku.edu/dspace/...ries.013op.pdf
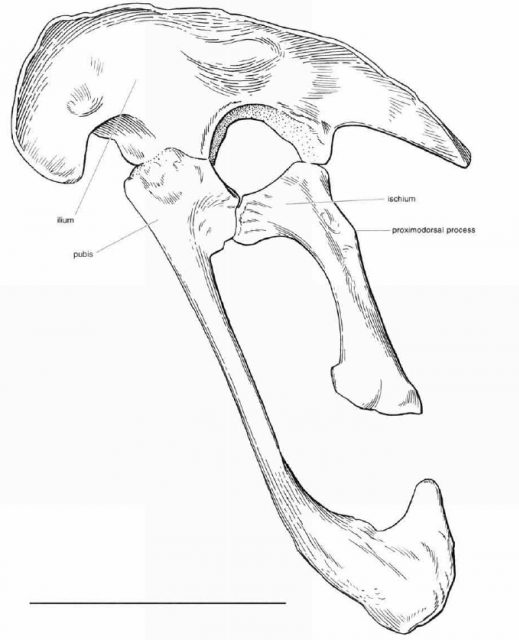
Hesperonychus:
http://www.pnas.org/content/106/13/5002.full

MODERN BIRD
Birds are the only living vertebrate whose hind limb includes three long bones in sequence. The innermost of the two long bones are similar to those found in most vertebrates. At the hip, the femur, is held more or less parallel to the ground and is bound to the hipbones by the massive thigh muscles. In effect, the femur is an addition to the hipbones and its rotation contributes little to the length of the bird’s stride.

During walking and running in birds, hindlimb movement is generated primarily at the knee and ankle joints; in humans, movement occurs at the knee, ankle and hip joints. The bird's thigh does not move substantially from its nearly horizontal position where it provides rigid lateral support to the thin walled air-sacs of the respiratory system. (Credit: Image courtesy of Oregon State University).
Note the pterosaur prepubis:
http://pterosaurheresies.wordpress.com/2011/11/06/whats-with-that-deep-prepubis/
The “dark wing” specimen of Rhamphorhynchus muensteri JME SOS 4785 (Tischlinger and Frey 2002) has one overlooked oddity worth mentioning. It had an incredibly deep prepubis (Figure 1.)
http://archosaurmusings.wordpress.com/2009/12/24/back-to-that-pterosaur-sacrum-pelvis
So much so normal, but pterosaurs also have a fourth pelvic bone in the form of the pre-pubis.http://www.bioone.org/doi/abs/10.1642/0004-8038%282007%29124%5B789%3ATAOBFA%5D2.0.CO%3B2?journalCode=tauk
The Auk 124(3):789–805, 2007
THE ANTITROCHANTER OF BIRDS: FORM AND FUNCTION IN BALANCE
Fritz Hertel1,3 and Kenneth E. Campbell, Jr.2
Abstract.—The antitrochanter is a uniquely avian osteological feature of the
pelvis that is located lateral to the postero-dorsal rim of the acetabulum. This feature
makes the avian hip joint unique among all vertebrates, living and fossil, in
that a significant portion of the femoral–pelvic articulation is located outside of the
acetabulum. This additional acetabular articulation occurs between the neck of the
femur and the antitrochanter, and operates as a hinge joint or ginglymus. It is complementary
to the articulation of the head of the femur with the acetabulum, which
is a pivot joint or trochoides. The size, location, and spatial orientation of the antitrochanter
were determined for 77 species of birds representing a variety of hindlimb
functions (e.g., highly cursorial, vertical clinging, foot-propelled diving) and spanning
a wide range of body sizes (swifts to rheas). The area of the antitrochanter is
a good predictor of body mass in birds; its position and orientation are reasonably
consistent within hindlimb morphofunctional groups, but not among all birds. The
antitrochanter serves as a brace to prevent abduction of the hindlimb and to absorb
stresses that would otherwise be placed on the head of the femur during bipedal
locomotion. The drum-in-trough-like form of the antitrochanter–femur articulation tends to assist in the transfer of long-axis rotational movements of the femur to the pelvis. The avian antitrochanter is a derived feature of birds that evolved as an aid in maintaining balance during bipedal terrestrial locomotion.
http://en.wikipedia.org/wiki/Pterosaur
Also:Pterosaurs' hip sockets are oriented facing slightly upwards, and the head of the femur (thigh bone) is only moderately inward facing, suggesting that pterosaurs had a semi-erect stance. It would have been possible to lift the thigh into a horizontal position during flight as gliding lizards do.
https://www.app.pan.pl/archive/published/app59/app20111109.pdf
Evolution of the pterosaur pelvis
https://en.wikipedia.org/wiki/Sacrum
The sacrum (/ˈsækrəm/ or /ˈseɪkrəm/; plural: sacra or sacrums;[1] Latin os sacrum[2]) in human anatomy is a large, triangular boneat the base of the spine, that forms by the fusing of sacral vertebrae S1–S5, between 18 and 30 years of age.[3]
In birds the sacral vertebrae are fused with the lumbar and some caudal and thoracic vertebrae to form a single structure called the synsacrum
https://en.wikipedia.org/wiki/Synsacrum
The synsacrum is a skeletal structure of birds and dinosaurs, in which the sacrum is extended by incorporation of additional fused or partially fused caudal or lumbar vertebrae.
http://www.prehistoriclife.xyz/history/mesozoic-birds.html
Archaeopteryx, as we have seen, had many features that are far from the condition found in living birds, including teeth, an unfused hand, a bony tail, no synsacrum, and gastralia.Subsequent events included the formation of a pygostyle as well as the development of the synsacrum and other features for a rigid trunk, all of which contribute to the efficient flight that characterizes modern birds.
Closer related yet to Aves were the toothed Ichthyornithiformes (see Figure 11.2). Unlike hesperornithiforms, ichthyornithiforms were excellent flyers (Figure 11.7). Ichthyornis, from the Late Cretaceous of North America, had a massive keeled sternum and an extremely large deltoid crest that was probably an adaptation for powerful flight musculature. In other respects, it shared many of the adaptations of modern birds including a shortened, fused trunk, a carpometacarpus, a pygostyle, a completely fused tarsometatarsus, and a synsacrum formed of 10 or more fused vertebrae. Found exclusively in marine deposits, ichthyornithiforms must have been rather like Mesozoic sea gulls - but with teeth.
http://www.reinhold-necker.de/seite14.html
The sacral enlargements of non-avian dinosaurs do not share much structural similarities with the lumbosacral specializations of modern birds. Furthermore these species were quadrupedal and had a long tail, i.e. there was no need for a sense organ of equilibrium. It seems that early birds with a pygostyle and a well- developped synsacrum show structural specializations which resemble that of modern birds. However, there is a need for more details and more examples of well- preserved fossils.
During evolution the reduction of the tail is accompanied by a melting of lumbosacral vertebrae named „synsacrum“.
http://bio.sunyorange.edu/updated2/pl%20new/55%20first%20birds.htm
Rahona ostromi lived in the late Cretaceous but its pelvic bones resemble those of the first birds. (Rahona means “menacing cloud.) It possessed a number of theropod characteristics (such as a pubic boot and the articulations between its vertebrae) but had the distinctive sickle claw and hyperextensible 2nd toe found in dinosaurs such as Velociraptor. It had primitive theropod characteristics such as a long tail, a saurischian vertebral articulation (also seen in Patagonykus), a pubic boot (as in Archaeopteryx and the enantiornithine birds) and some pelvic features (similar to Archaeopteryx and Unenlagia). It possessed a synsacrum which is an advanced feature (Forster, 1998; Gibbons, 1998).https://evodevojournal.biomedcentral.com/articles/10.1186/2041-9139-5-25
The oldest known bird, Archaeopteryx, dated to 150 million years ago, defines the clade Aves [5, 6, 7] or Avialae [8]. Its fully formed flight feathers, elongated wings, and evidence of capable powered flight, all ally Archaeopteryx with birds [9, 10]. Yet, the presence of teeth, clawed and unfused fingers, and an elongated, bony tail are characteristics shared with non-avian theropod dinosaurs. Paravians, including Archaeopteryx, are characterized by long tails [11, 12], some fusion of synsacral vertebrae, and varying flight capability (Figure 1). Most deinonychosaurians had between 20 and 30 caudal vertebrae. Oviraptorosaurs, probably the immediate outgroup to Paraves, had relatively shorter tails. These shorter tails were due not just to a modest decrease in the number of caudal vertebrae relative to other non-avian theropods, but more generally to a reduction in individual lengths of the more distal caudals [13]. Interestingly, several oviraptorosaurs have been documented to have the distal caudal vertebrae co-ossified into a pygostyle-like structure that braced a fan-like arrangement of retrices [13, 14, 15, 16]. Another more prominent independent reduction of tail length occurred in Epidexipteryx, a Mid- or Late-Jurassic maniraptoran dinosaur [17]. Its tail had only 16 caudal vertebrae with the distal ten tightly articulated to form a stiffened rod supporting four unique, ribbon-like, tail feathers.
https://static-content.springer.com/image/art%3A10.1186%2F2041-9139-5-25/MediaObjects/13227_2014_Article_146_Fig3_HTML.jpg
http://www.ivpp.ac.cn/qt/papers/201403/P020140314389417822583.pdf
The holotype specimen of Xiaotingia zhengi has completely closed neurocentral sutures on all exposed vertebrae and has a completely fused synsacrum, indicative of a late ontogenetic stage (probably adult).
Five sacral vertebrae form a short synsacrum (less than 60% as long as the ilium), as in other archaeopterygids and basal deinonychosaurs.
http://www.app.pan.pl/article/item/app20120093.html
W. Scott Persons, IV, Philip J. Currie, and Mark A. Norell
Oviraptorosaur caudal osteology is unique among theropods and is characterized by posteriorly persistent and exceptionally wide transverse processes, anteroposteriorly short centra, and a high degree of flexibility across the pre-pygostyle vertebral series. Three-dimensional digital muscle reconstructions reveal that, while oviraptorosaur tails were reduced in length relative to the tails of other theropods, they were muscularly robust. Despite overall caudal length reduction, the relative size of the M. caudofemoralis in most oviraptorosaurs was comparable with those of other non-avian theropods. The discovery of a second Nomingia specimen with a pygostyle confirms that the fused terminal vertebrae of the type specimen were not an abnormality. New evidence shows that pygostyles were also present in the oviraptorosaurs Citipati and Conchoraptor. Based on the observed osteological morphology and inferred muscle morphology, along with the recognition that many members of the group probably sported broad tail-feather fans, it is postulated that oviraptorosaur tails were uniquely adapted to serve as dynamic intraspecific display structures. Similarities, including a reduced vertebral series and a terminal pygostyle, between the tails of oviraptorosaurs and the tails of theropods widely accepted as basal members of the Avialae, appear to be convergences.
http://brantworks.com/resources/Allosaurus/ANTS_kit/ANTS-Allosaurus-Frank-Galef-detail-spine-BrantWorks.jpg

Feduccia
The (Archaeopteryx) synsacrum is also smaller and less fused, and the pubis is not fully reversed.
http://www.pterosaur.co.uk/PDB2012/P/index.htm
Pterosaurs all have a diapsid skull showing two openings behind the orbit. They have air sacs and pneumatic bones, a sclerotic ring around the eye and a distinctly fused synsacrum and pelvis.
The (Archaeopteryx) synsacrum consists of 5 fused vertebrae.
Subscribe to:
Posts (Atom)