Here is a
summary of the pterosaur to bird ideas of Larry Febo.
They are startlingly similar to what I have been proposing.
"NAME OF THEORY: Larry Febo`s , Birds are Pterosaur Offshoots
ORIGINATORS: May have been briefly considered by Seeley in the 1800`s. "Seeley (1881) and Weidershiem (1883, 1886) proposed a pterosaur-like ancestor for Archaeopteryx." (The Beginnings of Birds (1985) pg 16.) Otherwise, Larry believes he's the only one proposing this at the moment.
TIME OF ORIGIN OF BIRDS: Mid-Triassic (c 230mya).
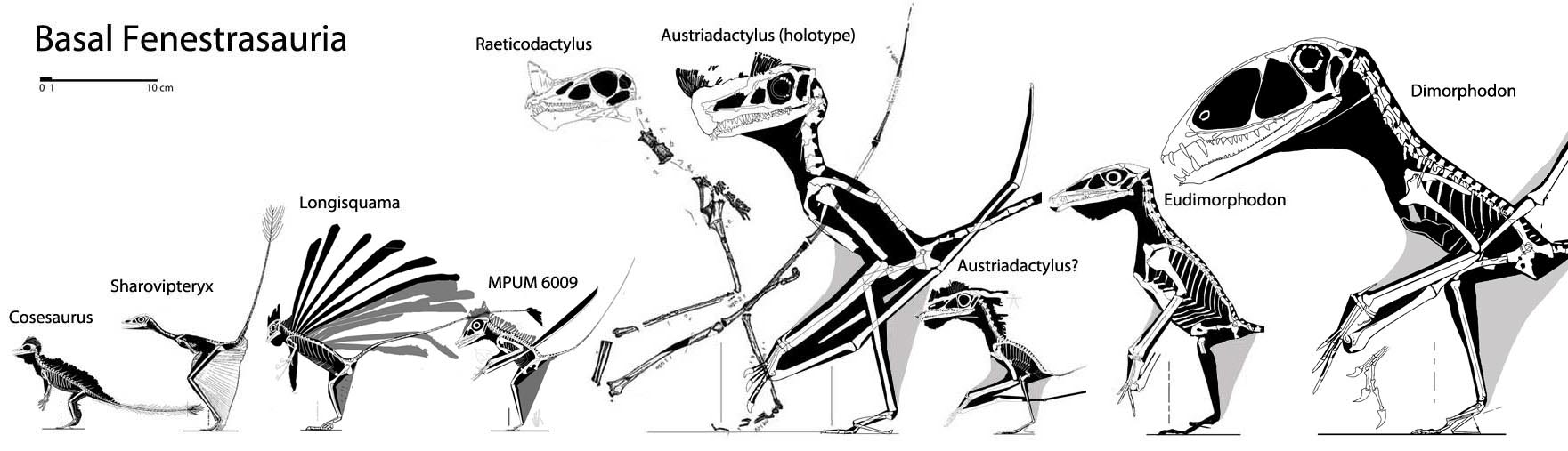
IMMEDIATE ANCESTOR OF BIRDS: A hypothetical Pterosaur-bird with small cranium, and relatively small wings, itself descended from the Prolacertiform group (with Cosesaurus as an appropriate example).
TIME OF FIRST FEATHER: Mid-Triassic
EXTINCT FEATHERED GROUPS: Protoavis (of the Triassic),...and of course the late Jurassic and early Cretaceous finds.
WEAKNESSES: Dependant on a hypothetical "missing link".
STRENGTHS: Resolves the "Digit Homology" controversy between the ornithologists and the paleontologists. The missing anterior digit is accounted for as being the pteroid bone, yeilding the homologous digit count 1,2,3 as preferred by paleontologists. Uses minimal dependence on the concept of Convergence.
PREDICTIONS: More pre-Archaeopteryx feathered birds will be found, eventually closing the gap back to the Mid-Triassic.
CHANGES NEEDED FOR CONVERSION TO 2F: Nothing simple would do it.
EVIDENCE REQUIRED FOR SUCH CHANGES:
GENERAL COMMENTS:"
OTHER RELATED MATERIAL:
http://dml.cmnh.org/1999Jan/msg00453.html
Hi folks, I just got around to reading "The Beginnings of Birds" (Hect et. al.). (Hey!...I told you I was playing "catchup" with my old paleontological interests!).Anyways, ...got to the article by Hinchliffe 'One, Two, Three' or 'Two,Three,Four', and it struck me immediately that my theory on "Birds as Pterosaur Descendants" seemed to fit in...(like that annoying missing piece of a large jigsaw puzzle that was never considered because of it`s "unlikely" color). The article describes how Holmgren (1955) and Montagna (1945) (quite a while back)..."claimed to find either a transient digit or its associated distal carpal preaxial to that developing digit which eventually forms the anterior digit of the adult wing". This transient element, I call "X", because I agree with the paleontological view that it cannot be called digit 1, as digits 1,2 and 3 are already identified (via phalangeal count), in both Theropod manus and Avian wing. Yet, it has to be something, as embryologists have determined its presence. ...There`s only one creature, (that I`m aware of), that fits the bill as having and "extra" preaxial "digit", and that would be the Pterosaur, with its "pteroid bone".
http://dml.cmnh.org/2001Jan/msg00135.html
From Larry Febo:
I`m not sure exactly how feathers developed,but actinofibrils arranged in a
pattern similar to flight feather rachis is very suggestive. Whatever the
path, a changeover from a fully functional membrane to feathers would have
to be gradual, and would require a "reason" for it to happen. I believe the
reason might have been the invasion of a new, and colder enviornment.
Perhaps a more upland enviornment, and natural selection acting on this
primitive wing to cut down on exposed area in a colder enviornment. Contour
feathers, once developed, could have extended to long flight feathers as the
membrane, and wing finger gradually (over generations) shrunk, until this
wing finger was itself no longer necessary.