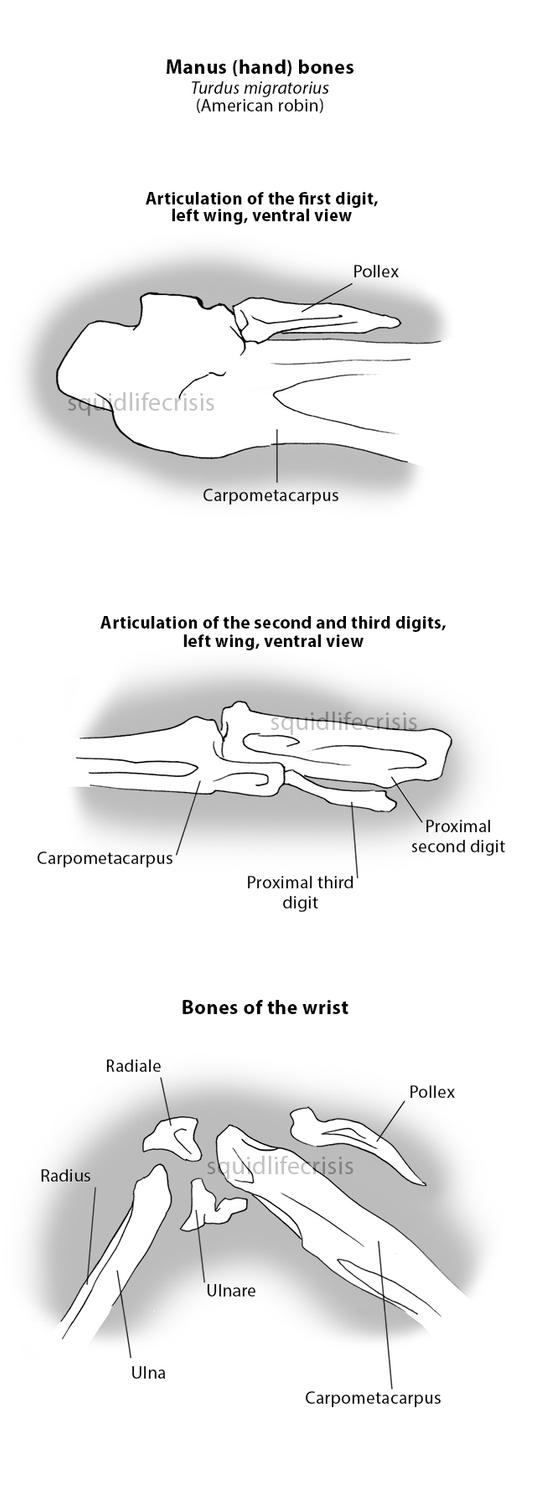
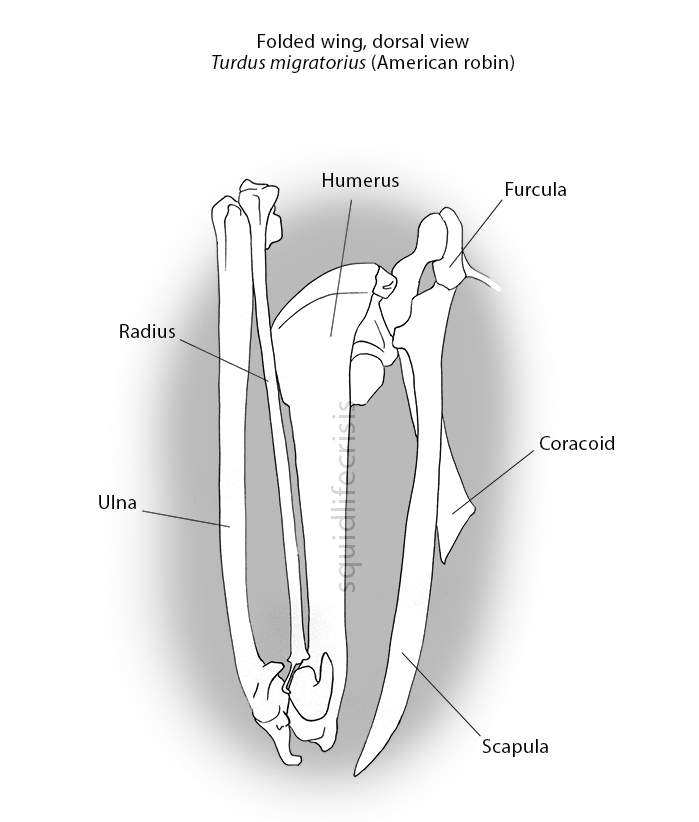
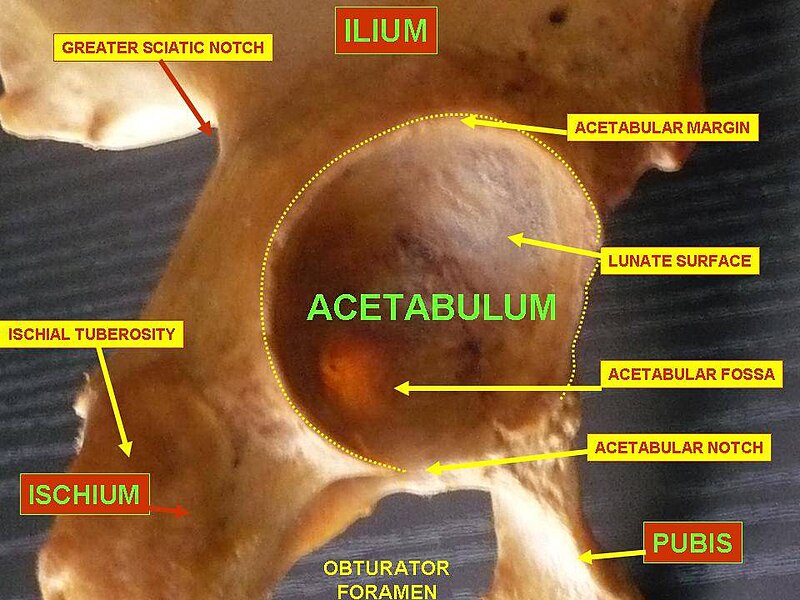
So the question arises:
How in the world could there be so many claims for years and years that birds evolved from dinosaurs?
In addition to the points noted above:
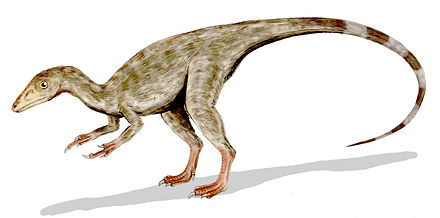
First, is the misleading convention of calling paraves "dinosaurs". So any bird-like character found in paraves is said to confirm the dino to bird theory. But paraves are not dinosaurs, they did not evolve from dinosaurs. People focus on the wrong place. The Achilles Heel of the dino to bird theory is that there is no connection between actual dinosaurs and paraves.
Next is to misinterpret the characters of actual dinosaurs as if they were bird-like or "proto" bird-like characters. Thus for example, we get the claim of "protofeathers" on ground-based coelurosaur dinosaurs, which does not stand up.
Also we get secondarily flightless paravians being called "non-paraves maniraptors" (eg. oviraptors). As if they were transitional between actual dinosaurs and arboreal paravians. That does not stand up. They are secondarily flightless members of paraves.
And also the cladistic analyses that have been done, generally include only dinosaurs and use an inappropriate outgroup. The very significant exception to this is the James and Pourtless study, which not co-incidentally found other explanations as credible as the dino to bird theory.