http://en.wikipedia.org/wiki/Hesperornis
http://en.wikipedia.org/wiki/Hesperornis"Hesperornis is an extinct genus of flightless aquatic birds that lived during the Santonian to Campanian sub-epochs of the Late Cretaceous (89–65 mya).
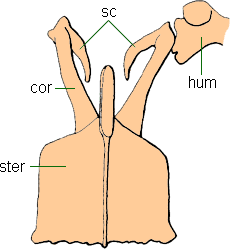
Although some of the smaller species might have been able to fly, Hesperornis and Baptornis had only vestigial wings. Like living foot-propelled diving birds, the femur and metatarsus were short, whereas the tibia was long. The legs were also set far back on the body, as in loons, grebes or penguins."
http://en.wikipedia.org/wiki/Grebe
"The cladistics vs. phenetics debate of the mid-20th century revived scientific interest in generalizing comparisons. As a consequence, the discredited grebe-loon link was discussed again. This even went as far as proposing monophyly for grebes, loons, and the toothed Hesperornithiformes "
http://www.jstor.org/pss/2413412
"A cladistic analysis of the skeletons of loons (Gaviidae), grebes (Podicipedidae), and the Cretaceous diving birds, Hesperornis and Baptornis, supports the hypothesis that they form a monophyletic group (here called the Gaviomorphae) within the class Aves. Two lineages within the gaviomorphs can be delineated: (1) loons + grebes, and (2) Hesperornis + Baptornis. The Early Cretaceous Enaliornis and the Late Cretaceous Neogaeornis are related to the second lineage; the Late Cretaceous Lonchodytes, often placed near loons, is apparently not a gaviomorph but perhaps a charadriiform. Skeletal evidence also suggests that penguins (Spheniscidae) are the sister-group of the Gaviomorphae. Arguments that similarities of gaviomorph taxa represent convergence are not well founded. First, morphological differences among taxa do not constitute valid evidence against their monophyly, as many previous workers have argued. Second, in no case has anyone supporting convergence presented a corroborated alternative phylogenetic hypothesis. A close relationship among gaviomorphs, penguins, and apparently also the Procellariiformes and Pelecaniformes implies that a lineage of aquatic birds was established very early in avian history, presumably in the Early Cretaceous or Late Jurassic." (Joel Cracraft 1982)
Looks like Joel Cracraft was on the right track!
(Note the issue here is not whether grebes and loons go together but that hesperornithiformes is linked to modern birds such as grebes and/or loons.)